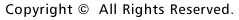|
画像をクリックすると拡大します。
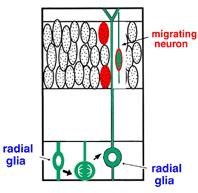
図1 大脳皮質における新生ニューロンの移動の旧模式図 |
画像をクリックすると拡大します。
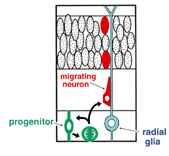
図2 RGCの分裂とニューロンの誕生。脳膜面側からDiIによって標識された1個のRGCの蛍光画像のタイムラプス記録。RGCは1.7-6.1時間の間にVZで細胞分裂し、2個の娘細胞を誕生。一方の娘細胞は親のradial glial process(赤矢印)を相続し、ついでapical process(緑矢印)の退縮にあわせて軟膜側へと移動しニューロンへと分化した。
画像をクリックすると拡大します。
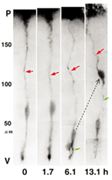
図3 RGCからニューロンとプロジェニターの産生。6-9.2時間の間にRGCから2つの娘細胞が誕生。一方は親のradial processを相続し(黒矢印)、ついでapical process(緑矢頭)を退縮して軟膜側へと移動しニューロンへと分化した。他方の娘細胞(青矢印)は、新規のbasal process(赤矢頭)を軟膜側へと伸長し、いったんその先端が境界面に達した後、再び細胞分裂して2つの孫娘細胞を誕生させた。
画像をクリックすると拡大します。
図4 ニューロン誕生と移動の新模式図 |
|
はじめに
大脳皮質は脳膜面に並行した6層の細胞層から構成され、層ごとにタイプ(形態と機能)の異なるニューロンが配置されています。この整然としたニューロンの配置が大脳の生理機能の基盤であることは、種々のマウスミュータントやヒトの先天性脳神経疾患における所見から明らかであります。大脳の構造の形成原理を理解することが、ヒトの疾患の病理解明のためにも、また、再生医療への歩みの一つとしても重要です。私たちのチームでは、大脳皮質における皮質ニューロンの誕生、移動ならびに6層構造の形成機構を、細胞/組織培養法などを利用して研究しています。
皮質ニューロンの誕生と移動
大脳皮質においてニューロンがどのように誕生し、そして移動して分化するのでしょうか。1970年代にRakicによって提唱されたradial glial guidance 説は、ニューロン誕生と同時に起こるその移動を説明する概念として、多くの神経生物学者に受け入れられてきました。その基本骨子は、脳室帯(ventricular zone, VZ)に遊離細胞として誕生したニューロンは、隣接して存在する放射状グリア(radial glial cells, RGCs)に接触し、ついでRGCsの放射状突起(radial glial process)に沿って脳膜側へと移動するというものです(図1)。RGCsは、その細胞体がVZにあって、これから脳膜面に達する長いradial glial process(rgp)と内腔面に達する短い突起(apical process)でもって大脳壁を縦断し、またVZで皮質ニューロンが産生される期間、細胞増埴を停止している静的な細胞とされてきました。私たちは胎生期(E13ないしE14)のマウス大脳を用い、微小なDiI結晶で脳膜面からRGCsを標識し、ついで独自に開発した3次元スライス培養法を用いて、蛍光顕微鏡下に生きたRGCsの細胞行動をリアルタイムないしタイムラプスに観察記録しました。その結果、まずRGCsは静的な細胞でなく、細胞分裂を起こしてニューロンおよびプロゲニター(progenitor,P)を産生することが示されました。すなわち、RGCsは神経上皮細胞に属する細胞であることが判りました。ついで、その細胞分裂に際して、親のrgpが消滅することなく一方の子細胞に相続されることが示されました(図2)。親のrgpを相続して誕生したニューロンは、当初rgp内を細胞核が脳膜側へと転位(nuclear translocation)し、ついで、apical processの脳室面からの離脱、rgpの短縮にともなって脳膜側へと移動することが示されました。一方、RGCの分裂によって誕生したP細胞は、すぐさまbasal processを伸長し、それが脳膜面に到達した後、つぎの細胞分裂を起こして孫細胞を誕生することがわかりました(図3)。これらの観察は、これまで細胞系譜が異なるとされてきた新生ニューロンとRGCが、実は同じ系譜のものであることを示し、RGCの特異な細胞形態を維持し、それが固有する自律的な遺伝子情報に従ってニューロンが誕生することを示唆しています。皮質ニューロンの誕生と移動に関するこのような新たな概念と意義については、最近のCerebral CortexおとびGlia誌にそれぞれ特集号として編集されていますので、興味を抱かれる方は参考にしてください。このように大脳壁組織の3次元培養法を開発することによって、皮質ニューロン誕生の生のドラマをはじめて観察することができ、またニューロン誕生の新たな概念の誕生をみることができました。
大脳皮質の6層構造の形成機構
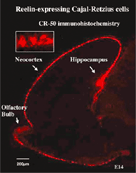
大脳新皮質は6層の細胞層から構成され、層ごとにタイプ(形態と機能)がことなるニューロンが配置されています。同じ層のニューロンは同時期にVZに誕生し、早生まれのものがより深層に、遅生まれのものほどより表層に配置されます(inside-out)。私たちは御子柴研などとの共同研究を通して、皮質に最初に現れ、辺縁帯(marginal zone)に位置するCajal-Retzius 細胞から特異的に分泌されるReelinによって、皮質ニューロンの空間配列が制御されていることを明らかにしました(図5)。ミュータントマウスのリーラー(reeler)では、この分子が欠落していることによって、6層構造が異常になります(outside-in)。また移動する皮質ニューロンが、apoER2, VLDLR, CNR等のReelin受容体を通してこのシグナルを受容し、細胞内アダプター分子のDab1のチロシンリン酸化を一過性に促すことが明らかにされてきました。Reelin欠損マウス(reeler)、apoER2とVLDLRの2重欠損マウス、Dab1欠損マウス(yotari)、さらにDab1のチロシン残基をフェニルアラニンに置換したマウスにおいては、ほぼ類似した層構造の異常がみられます。しかし、このリーリンシグナル伝達が細胞のモーター分子といかに連携してニューロンの動きに関わっているかは明らかでありません。ところで、皮質ニューロンがinside-outに配置されることは、遅生まれのニューロンが先に生まれたニューロンよりもより表層に移動することになります。しかしまだ誰も、生きた組織で、遅生まれのニューロンが早生まれのニューロンを追いこして移動することを観察記録することに成功していません。わたしたちは、誕生日の異なる移動するニューロンをそれぞれ別の蛍光タンパク質で標識し、より改良された3次元培養法を用いて、このようなニューロンのダイナミックな移動を記録することを試みています。
ところで、最近に報告されたヒトの常染色体劣性遺伝性リーリン病では、小脳の形成不全に加えて、大脳がlissencephalyになることが示されています。げっ歯類の大脳はもともと滑面脳ですので、大脳がlissencephalyになるとはreeler マウスから予想されないことでした。reelerマウスの小脳では、葉構造が形成されません。ラボでは、葉構造形成におけるReelinの役割を手がかりに、大脳における“しわ”構造形成のメカニズムも調べています。
画像をクリックすると拡大します。
図5 Cajal-Retzius細胞。Cajal-Retzius細胞は外套(pallium)に最初に現れるニューロンで、辺縁帯に配置する。この細胞が特異的に産生、分泌するReelin分子によって、皮質ニューロンの層配列が制御されている。 |
|
おわりに
私たちは脳の組織構築の原理を明らかにするうえで有効な、細胞/組織培養実験系をこれまでにいくつか開発してきました。本稿では脳のスライス培養をとおして見い出した、大脳皮質におけるニューロン誕生と移動、ならびに層形成のメカニズムについて紹介しました。その他にも、ニューロンとグリア細胞の細胞系譜の解析に有効な低密度培養法(low cell density culture)の開発、皮質ニューロン間の細胞接着の解析に有効な再凝集培養法(reaggregation culture)の開発、小脳皮質形成の解析に有効な胎生期の小脳の器官培養法(organ culture)の開発などを行ってきました。これらの実験手技は国内外の諸研究機関で試され、いくつかの成果を生み出してきました。このように、他のラボの研究支援になるような実験手技の開発も、私たちのラボの重要な使命と考えています。 |
|
| |
 |
|
|