






Our visual system normally provides us with reliable, unambiguous information about the external world. Yet there are also numerous situations where visual perception misleads us or provides us with ambiguous results. Ambiguous patterns are amongst the most widely known phenomena in psychological research of the visual system. When, for example, we are looking at Attneave's triangles (Figure 1), at first all seem to point in one particular direction. But after a while, they start to point in another direction. And so on, indefinitely.
Often it has been assumed that such switches are an all-or-none phenomenon. Observe, for instance, that when the triangles change pointing direction they do so all at the same time. But as Attneave already pointed out, this is not always the case; sometimes the triangles lose their sense of direction or point in inconsistent directions, leaving a weakened image of the whole. Similar processes also occur for other ambiguous patterns, such as the widely known Necker-cube, although they are somewhat harder to demonstrate. Global consistency is not as rigidly maintained by the visual system as we might have expected.
All-or-none or partial, the collapse of global perceptual structure plays an important role in the occurrence of a reversal. This has, indeed, been the assumption for a number of neural theories that describe the switching phenomenon. The Gestalt psychologist Wolfgang Kohler thought that the collapse was due to neural fatigue. Intrinsic instability of neural synchronization is now often believed to be the cause of the collapse of the pattern. All these notions rest on the assumption that there is a unitary, low-level brain event underlying the switching phenomenon. Our recent research, however, has shown that in fact a cascade of events is involved in the breakdown of a given structure and its replacement by another one.
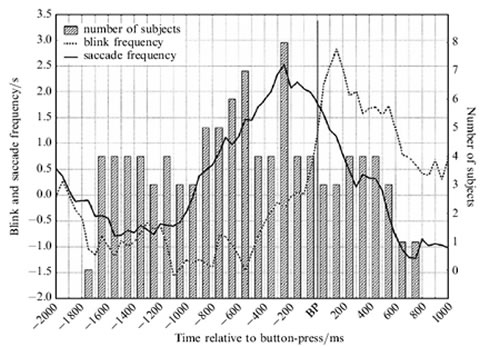
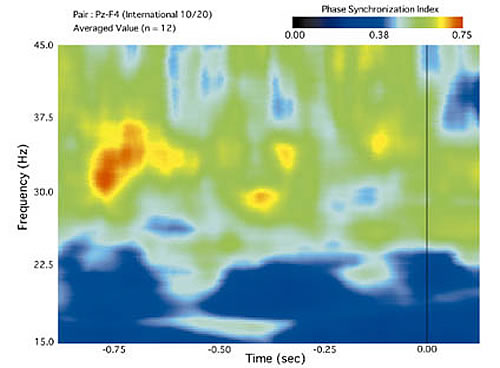
Figure 2, illustrates the sequence of events prior to the switching response, in eye-movement and EEG respectively. In Figure 2A, participants were asked to press a button (BP), any time they see the entire set of triangles change pointing direction. We registered the blink (closing the eyes and opening them again) and saccade (a sudden eye-movement from one eye-fixation position to another) frequency, and pointing-consistent eye-movement (intervals during which participants moved their eyes significantly more frequently in the pointing direction of the triangle indicated by the button press than in any other direction). From 1700 ms to 700 ms prior to the switch response, blinks and saccades are suppressed. In the literature, these phenomena are understood as correlates of cognitive activity. We may, therefore, infer that cognitive activity initiates the switching response. After this period, from approximately 700 ms prior to the button press, we observe an increase in eye-movements that are consistent to the new pointing direction indicated by the button-press. These eye-movements can be associated with conscious awareness of the new pointing direction. After the button press the number of blinks is larger than normal, apparently a form of compensation.
In Figure 2B, we show the time-frequency plot of synchronization in EEG activity for switching in the Necker cube. Warmer colors indicate a higher degree of synchronization between a parietal and frontal site. In this plot, there are three brief events marked by high coherence. The first, around 700 ms, corresponds to the moment where in Figure 2A saccade frequency starts to increase again, and eye-movements are becoming consistent with the new interpretation. This indicates that the new pattern is being established. The second interval of increased synchrony, 350 ms prior to the response, corresponds to the maximum in consistent eye-movements. It may, therefore, indicate the capture of attention by the new pattern. The final bit of synchronization occurs just 120 ms prior to the switch. This may correspond to the manual response.
Figure 2 indicates that switching involves a cognitive event, followed by a sensory event, followed by a motor event. This cascade of events involves all cortical functions in a top-down fashion. This raises the interesting question, whether we can exert voluntary control on our switching behavior. When instructed to try not to switch, switches will become more rare (although care must be taken to distinguish this from merely being more cautious in reporting the occurrence of a switch). When asked to hold one of alternative interpretations, participants can successfully do so, to the extent that that interpretation is more likely to occur over a time interval. But this will not prevent them from switching to another one. Control, therefore, is only possible to a limited degree.
Perceptual switching provides us with an important illustration that the visual system is, in fact, an active system. It doesn't always wait for new information to arrive before assuming a new state, but sometimes changes its state by itself. The results presented suggest that switching is initiated from the higher-order visual functions and is not a result of instabilities within the early visual areas alone. In this sense, there is a striking relationship with mental imagery, which remains to be explored further.
