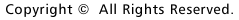|
 |
|
 |
学習と記憶における海馬のNMDAレセプターの役割
|
RIKEN-MIT脳科学研究センター
グループディレクター
利根川 進 |
|
|
| |
|
画像をクリックすると拡大します。
図1.リコンビネースCreトランスジェニックマウス(T29-1)とlac2トランスジェニックレポーターマウスの掛け合わせにより作られたマウスの、脳をX-gal染色した写真。円弧内の青色はCre/loxPの欠損の組換えが海馬のCA1領域に特定して生じたことを示す。
|
私は、いわゆる心と脳の問題に常に興味を抱いてきた。脳を研究することにより、どれほど心を理解できるのだろうか。私の主な研究人生での経験は分子生物学、免疫学であって、神経科学についてはほとんど何も知らなかった。神経科学への一般的な興味が本物の興味へと変わったのは、1980年代の終わりにアルシノ・シルヴァが私の研究室にポストドクとして入ってきてからである。当時、私の研究室では免疫学の研究を行っており、我々は遺伝改変マウスを作っていた。アルシノと研究課題の可能性を話していたとき、彼も神経科学に興味を持っていることがわかった。私の知る限り、それまで誰も強力な遺伝子工学の技法を神経科学に応用したことはなかった。
認知現象にともなう脳内で起こる事柄を解読するには、生きた動物をそのまま使用する実験方法を取り入れなくてはならない。ノックアウトマウス(遺伝子改変マウス)は、非常に強力な道具となることは明らかに思えた。ノックアウトマウスでは、特定の遺伝子が取り除かれている。したがって、通常のマウスと比べてこれらのマウスに見られる認知や行動の欠陥は、直接的または間接的に1つの遺伝子の欠損に結び付けられる。同様な方法は無脊椎動物の行動を分子的に研究するのに他の人々によって用いられたが、脊椎動物には応用されなかったし、ましてや哺乳類に適用されたことはなかった。
画像をクリックすると拡大します。
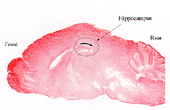
図2.写真は、CA1限定NR1ノックアウトマウスにおいて、NMDAレセプター関連シナプス可塑性(LTPとLTD)がCA1のシナプスでは障害されているが、歯状回(DG)または体性感覚野(CC)のシナプスは障害を受けていないことを示す。T29-1とWTはそれぞれCreトランスジェニックマウスと野生型マウスである。またfNR1マウスは、NR1遺伝子の上流および下流に一対のLoxPサイトが挿入されたマウス。これらを対照マウスとして用いた。
|
我々は、認知機能として学習や記憶を研究することにし、ノックアウトの標的としてCaMKIIサブタイプを選択した。1992年にこの研究が神経科学での最初のノックアウトマウスの報告となった。α-CaMKII遺伝子の欠損は、シェファー側枝CA1シナプスでの長期増強(LTP)の欠損と空間学習の障害を発生させた。しかし本件の発表以前に、従来形ノックアウトマウスを用いたこの方法には限界があることに気がついていた。この限界とは、我々が興味を持っている遺伝子が、動物の組織全体でまたその一生を通して取り除かれているからである。CaMKIIαノックアウトマウスでは、発生学的には明らかではない欠陥は見られなかったが、もっと細かい欠陥を排除できなかった。問題とされている蛋白(この場合CaMKIIα)が全体的に欠損していることで、CA1LTPと空間学習の因果関係を成立させえなかった。
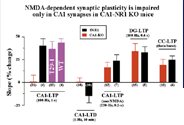
1993年に、もう一人のポストドク、ジョー ・シエンが私の研究室に着任した。我々は、特定の遺伝子の欠損を脳の一部に限定する第2世代のノックアウト技術の開発を始めた。2年間の苦労の結果、ジョーは遺伝子の変化が前脳部の特定の神経に限定される数種のマウスを作り出した。このマウスの一系統で遺伝子の変化が海馬のCA1領域に限られていることを知り、我々は興奮した。ジョーはさらに、この系統のマウスを用い、NMDAレセプター遺伝子であるNR1の欠損がCA1の錐体細胞に限られているマウスを作り出した(図1)。ジョーともう1人のポストドク、パト・ファルタは、これらのマウスではLTPやLTDはシェファー側枝CA1シナプスでは欠損しているが、穿通枝路歯状顆粒細胞シナプス(perforant path dentate granular cell synapses)等の他の場所では正常であることを示した(図2)。彼らはまた、突然変異マウスでは、海馬に関連の学習が欠損していることを発見した(図3)。さらに、廊下の反対側のマシュー・ウィルソンの研究室との共同研究の結果、これらの突然変異動物は、正常な場所野をCA1につくれないことを証明した(図4)。これらすべてのデータは記憶がシナプスを基にして行われるとしたヘブの仮設を強力にサポートする証拠となった。

画像をクリックすると拡大します。
図3.モーリス水迷路探索試験実験中の、正常マウスと(CA1限定NR1)ノックアウトマウスの三次元グラフ。対照マウスは捜索を訓練された場所を集中的に(そこには訓練中にはプラットホームがあった)、遺伝子変異マウスは全ての迷路を平等な割合で捜索した。
|
私はアルシノに、我々はNR1遺伝子のノックアウトをCA1領域に特定する技術を開発したようだと伝え、いつも明るく熱心なアルシノは「おお、すごい、おめでとう」と叫んだ。CA1は記憶形成に関係するとされた海馬の中で主要な解剖学上の領域でありシェファー側枝CA1シナプスはまさに神の授け物と考えられる。
遺伝子操作を脳の他の特定領域や他のタイプの細胞に対して行う技術を開発できるのであろうか。最近の我々の研究によれば、これはまさに可能である。例えば、我々の研究室の他のポスドク中沢一俊は、発生後の海馬のCA3錐体細胞をノックアウトする方法を開発することに成功した。この方法を用い、彼はCA3限定NR1ノックアウトマウスを作った。CA1限定NR1ノックアウトマウスに対し、この新しい変異マウスは海馬関連学習面では正常である。しかし、このマウスでは記憶想起で著しい障害がある。CA3限定NR1ノックアウトマウスで作られた場所細胞の特性は、行動レベルで見られた発見とうまく一致した。我々は、同じグルタミン酸・レセプター(つまり、NMDAレセプター)は、それがどのタイプの海馬の細胞に発現されているかによって、記憶の過程で大変異なる役目を果たしていると結論づけることができる。学習や記憶の研究、ひいては他の認知機能の研究における領域限定ノックアウトマウスの重要性は、これらの研究により実証された。

画像をクリックすると拡大します。
図4.すべての行動環境において、CA1限定NR1ノックアウトマウスの場所領域はかなり大きかった。それぞれの行動環境における対照動物の2つの錐体細胞とノックアウト動物の2つの錐体細胞の場所特異的活動度の図。それぞれの図の最高占拠量が最高活動量を示すように調整されている。 NMDAR1 CA1-ノックアウト動物の錐体細胞のフィールドの大きさは線形軌道(1次元)環境とオープンフィールド(2次元)環境で有意に大きかった。
|
さらに最近、この領域限定に組み合わせて、時間に対して可逆的に遺伝子産物(酵素や受容体)の機能を制御する遺伝子操作法が開発されつつある。この技術は、げっ歯類による認知や行動の基盤になっている脳の機構の解明に大いに役立つであろう。 |
|
|
|
 |
|
|