研究室のメンバー。左から順に:坂井理映子、松川浩、アレクサンダー・ウォルフ、高田綾子、松下真一、トーマス・クヌッフェル、ディネッシュ・ラジ、ウォルター・アケマン、栃谷史郎。撮影時不在メンバー:ヴィクトリア・コティンノ、高橋泰城
神経回路ダイナミクス:ニューロンの社会学
神経回路メカニズム研究グループ
神経回路ダイナミクス研究チーム
チームリーダー
はじめに
小さな細胞の集合体である脳は、神経細胞間のシナプス結合を介してコミュニケーションを行っている。神経細胞(ニューロン)では情報の送受信のみでなく、さまざまな情報を統合し判断するという機能も備えている。
20世紀初頭、
Santiago Ramo y Cajal
がニューロン説を提唱して以来、神経学者によってニューロンの巧妙なコミュニケーション能力が発見、報告されてきた。しかし今日においても、個々のニューロンの活動は十分に理解されたとは言えず、例えば細胞間の情報交換や統合がどのように調整されているのかなど解明されていないことも多い。また仮に単一ニューロンの活動が把握できたとしても、集合体としての機能は個々のニューロンの活動からは予測しきれないものである。脳全体の機能を研究することは「ニューロン社会学」であり、グループごとの行動パターンや相互作用による全体への影響などを考慮する必要がある。そもそも「社会学」というのは人間の社会的共同生活の構造や機能について研究する学問であるが、脳の仕組みを研究することはまさしく「社会学」であろう。つまり我々の脳の研究においても人間の社会学を研究するのと同様に、適した実験的方法論とそれによって得られた結果 を適切に解析するための理論的骨格が必要となるのである。
画像をクリックすると拡大します。
図1 本研究室における光学的手法を用いた研究の発展。従来からの微小電極による電位 測定に光学的イメージング法を併用し、カタツムリのニューロンの活動電位(A、1984年)や単一の培養プルキンエ細胞(B、1989年)、小脳切片培養標本でのプルキンエ細胞(C、1990年)の電位 変化を記録している。
実験的方法論
それでは、どのようにすれば多数のニューロンの相互作用を観察することができるだろうか?社会学者であれば、ある一集団の活動を観察することで、各個人の行動が意味するもの、そしてその背景にあるものを理解しようとするであろう。脳においても、このような「集団力学」をニューロン間のコミュニケーションによって生じる電気的信号(膜電位 の変化)を観察することによって把握することができよう。なぜなら、個々のニューロンから送信されたメッセージは活動電位 として、また受信されたメッセージはシナプス後電位として記録することができるからである。しかし実際には多数のニューロンが同時にメッセージを送信するので、我々は同時に多数のニューロン間で行われる情報交換、処理過程を記録することが必要となる。ここで問題となるのは、どのようにすれば同時に多数のニューロンから発せられる電気的信号を記録できるかということである。小さなニューロンは脳内で三次元に詰め込まれているため、すべての細胞に配線を施し
画像をクリックすると拡大します。
図2 小脳および大脳皮質体性感覚野におけるニューロン活動の光学的イメージング。
左:小脳表面への電気刺激によって小脳平行線維束の興奮が誘起され、シナプス後電位 のシグナル変化として捉えられている。
右:体性感覚野の第4層以下を電気刺激することで、2つのバレルに興奮が観察される。
て電気的信号を検出することは難しいが、光学的イメージング技術の開発がこれを可能にした。たとえば、膜電位 の変化に伴ってその蛍光を変化させる色素でニューロン膜を染色すると、同時に多数のニューロンの電位 変化が蛍光の変化として記録できる。私自身も長年、このような電位感受性蛍光色素(VSD)を用いてさまざまな実験を行ってきたが(図1)、さらに近年では、高速イメージング機器の発達によって哺乳類の脳切片における電気活動をも記録することが可能となった(図2)。図2に示すようにVSDと高速イメージングシステムを用いることにより、大脳や小脳での活動電位 伝播の様子や、それに続くシナプス後電位の変化を高時空解像度で視覚化することができるのであ
る。
画像をクリックすると拡大します。
図3 電位感受性蛍光蛋白質(原型、VSFP)を発現したHEK293培養細胞。
左:透過型顕微鏡写真
右:蛍光イメージ。VSFPを発現した細胞(左図矢印)の細胞膜は明るい蛍光を示すが、VSFPを発現していない細胞(左図*印)は蛍光を示さない。
このようにVSDは有用ではあるが、一方でその実質感度の低さや毒性の可能性、また目的とする特定の細胞のみを染色することができないため、実際にどの細胞が蛍光変化を示しているのか明確には判定できないなどの限界がある。そこで我々はこれらVSDの短所を克服できるような膜電位 感受性蛍光蛋白(VSFP)の開発に着手したわけである。我々が開発したVSFPは、膜電位 依存性カリウムチャンネルの電位感受性領域に変異型緑色蛍光蛋白を組み込んだもので、単一細胞レベルでの小さな電位 変化を感知し、またミリ秒の速さの電位変化にも追随することができる。またVSFPは目的とする特定の細胞にのみ発現させることもできるため、VSDを用いた場合よりも高いシグナル/ノイズ比を得られるのである(図3)。次のステップは当然のことながら、このVSFPを特定の脳細胞に発現させたトランスジェニックアニマルを作成することである。すでに我々はニューロン特異的プロモーターを用いpHおよびクロールイオン濃度感受性変異型緑色蛍光蛋白を導入したトランスジェニックマウスの作成に成功している(図4)。
理論的骨格
図4 海馬CA1領域の錐体細胞に発現したpHおよびクロールイオン感受性変異型緑色蛍光蛋白。s.p.:錐体層、s.r.:放射状層。
脳のコミュニケーションを理解するためには、まずそこで使われているいわゆる「言葉」を理解しなければならない。ニューロンからの送信メッセージを例にとれば、「ニューロン語」として重要なのはニューロンの平均発火率(rate-coding hypothesis)なのか、あるいはいかに適時に発火が起こること(temporal-coding hypothesis)なのかが争点となるであろう。temporal-codingが重要であることはニューロン間に相互作用があることを考慮すれば自ずと明らかである。社会学の立場から言うと「それぞれの個人が何をどれだけ言うかはもちろん大切なことであるが、その重要性はいつ、誰が発言し、誰が聞きどのように応答したかによって左右される」ということである。つまりニューロン間の相互作用によって「発言」が真の「情報」となるのである。ニューロンの情報処理過程ではグループ内でこれらの情報の統合や変換も行われ、導かれた結論が行動や認識に直接関与していることもある。人間社会においても共同声明が個々の意見よりもはるかに内容が充実しているのと同じである。また最近、ニューロン活動の振動(oscillation)と同期(synchronization)という現象に注目が寄せられている。これらの現象は temporal-codingのためのリズムとりの役目を担っているのか、あるいは単にニューロンの情報処理過程を反映しているものなのかわかっていない。
人間社会は外的要因によって変遷を繰り返すが、それはニューロン社会でも同様である。たとえば感覚組織に見られるように、外界からの刺激はニューロンで「ニューロン語」に翻訳され、その情報をもとに経験が生かされた適切な判断が下されている。ニューロン社会も記憶と学習によって幾多の変遷を経ているのである。
これから
我々には2つの大きな目標がある。一つは記憶と学習に関連して、新しい情報という刺激に呼応してニューロン間の相互作用にどのような変化が起こるのかを詳細に検討することである。一般 的には短期記憶はシナプスレベルで起こり、この変化はシナプス伝導の変化として観察されるが、長期にわたる記憶には発達過程の脳に見られるような情報処理システムの再構築が必要とされている。このような記憶学習過程に伴うニューロンの活動を光学的に可視化できればすばらしいことであろう。二つ目は、ニューロン活動の振動(oscillation)と同期(synchronization)に関することである。synchronizationは分散された情報の統合処理に関与しているという仮説があるが、光学的イメージング技術を応用すればこれらの現象の生物物理学的仕組みやその特徴が明らかになるかもしれない。このように我々が開発した光学的イメージング技術を用いての研究は、集合体としてのニューロンの役割に関する新しい理論的概念を展開する可能性を秘めているのである。






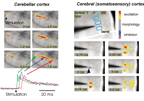
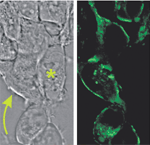
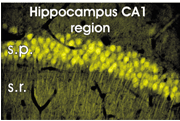 図4
海馬CA1領域の錐体細胞に発現したpHおよびクロールイオン感受性変異型緑色蛍光蛋白。s.p.:錐体層、s.r.:放射状層。
図4
海馬CA1領域の錐体細胞に発現したpHおよびクロールイオン感受性変異型緑色蛍光蛋白。s.p.:錐体層、s.r.:放射状層。