|
はじめに
私たちが視覚的に外界を認識するとき、脳はどのように働いているのでしょうか。本研究チームでは、脳の情報処理能力とそれを支える神経回路網の形成原理を理解するために、哺乳類の大脳皮質第一次視覚野に見られるコラム(方位
コラム、眼優位コラム、運動方向コラムなど)と呼ばれる機能的構造に注目して、理論と実験の両面
から研究を行っています。
私たちは、光計測法を用いて様々なコラム構造の2次元パターンを観察し、コラム相互の関係や情報表現様式を調べていますが、同時に、私たちが開発した「神経活動に依存する自己組織化の数理モデル」を用いてコンピュータシミュレーションを行い、実験で得られたコラム構造を再現することによって視覚野のコラム形成における視覚経験の役割について検討しています。

画像をクリックすると拡大します。
図1 運動方向コラムの画像処理
18野の光計測実験によって得られた生の運動方向コラムの2次元パターン(上段)とノイズ成分を取り除いた運動方向コラムの2次元パターン(下段)
|
光計測データの解析法
哺乳類の大脳皮質視覚野には様々な視覚特徴量
を表現するコラム構造が存在し、視覚情報処理を行うための機能的な参照地図を与えるものと考えられています。そこで、視覚情報の脳内表現と皮質神経回路による視覚情報処理のメカニズムについて理解するために、光計測法を用いてコラム構造のイメージングを行っています。私たちが用いている光計測法は、内因性シグナル計測法と呼ばれ、神経活動に伴う代謝活動の変化や形態変化を脳組織からの反射光の変化として捕らえるものです。
しかしながら、この計測法は微弱なシグナルから神経活動に関連する成分を計測するものなので、計測された画像データを適切に処理する必要があります。計測された画像データを詳細に調べたところ、コラムのイメージを劣化させている主なノイズ成分は、空間的にゆるやかに変化する刺激に依存しない成分であることがわかりました。そこで私たちは、そのノイズ成分を直行多項式を用いて抽出し、計測されたシグナルから差し引くことによって、刺激に依存するシグナルを効率よく求める方法を開発しました。図1に示すように、この方法を用いることによって、従来ノイズに埋もれて計測できなかったコラム構造も捕らえることが可能になりました。
|
|
神経活動に依存する自己組織化のモデル
では、コラム構造はどのようにして発達脳において形成されるのでしょうか。私たちは、この疑問に数理モデルを用いてアプローチしています。視床の中継核である外側膝状体にあるニューロンでは、明暗の光刺激に反応する視野上の領域(受容野)が同心円上に構成されているのに対して、視覚野ニューロンでは、明暗の光刺激に反応する領域がある傾きをもって平行に並んでいます。その結果
、明暗の縞を視野に提示すると特定の傾き(方位)に選択的に反応するわけです。また視覚野では、近傍のニューロンの受容野特性が類似しているために、方位
や運動方向のコラムが秩序だった配列を呈するのです。
こうした視覚野に特徴的な受容野の形成と方位
や運動方向のコラムの形成は、私たちが開発してきた「神経活動に依存する自己組織化の数理モデル」によって再現できることがわかってきました。このモデルは、外側膝状体のニューロンから視覚野のニューロンへのシナプス結合は、外側膝状体ニューロンのスパイク活動と視覚野ニューロンの電位
上昇が同時生起したときに強化される、とういうヘッブ仮説に基づく理論です。 図2は、様々な傾きの縞模様を提示しながら自己組織化のシミュレーションを行ったときに得られたモデル視覚野ニューロンの時空間受容野の典型例を示しています。また、図3は、同じシミュレーションによって得られた方位
コラムパターンを示しています。
|
|

図2
シミュレーションによって得られたモデル視覚野ニューロンの時空間受容野のスナップショット
赤と緑の領域は、スポット光を照射したときにオンまたはオフ反応を示す受容野内の部分領域を表します。
|
-- |
|

画像をクリックすると拡大します。
図3
シミュレーションによって得られたモデル視覚野の方位コラムパターン
|
奥行き運動方向のコラム
さて、従来光計測によってイメージングされてきた方位
コラムや眼優位コラムは基本的には視床からのフィードフォワードの神経連絡によって決定されると考えられていますが、コラムによる視覚情報表現がすべて視覚野へのシナプス結合のパターンによって決まっているとするならば、大脳皮質に存在する多くの皮質内神経連絡は視覚情報処理においてどのような役割を担っているのであろうか、という疑問が生じます。
このような疑問に対する答えを得るために、私たちは「奥行き運動方向」に関する情報の視覚野における表現様式を光計測法を用いて調べてみました。物体の奥行き方向の運動は、網膜に投影された像のサイズ変化や、左眼と右眼での網膜像の運動方向の相違に基づいて知覚されるものと考えられています。私たちは、特に後者のメカニズムに注目し、奥行き方向に運動する視覚刺激を、液晶シャッターによって開閉するゴーグルを用いて実現しました。
ディスプレイの画面上に提示した反対方向に移動する縦縞を120Hzで左眼と右眼に交互に見せることによって、縦縞が奥行き方向に運動するように感じさせることができます。例えば、左眼には右方向に移動する縞を、右眼には左方向に移動する縞を見せると、観察者は縦縞が近づいて来るように感じます。逆に、左眼には左方向に移動する縞を、右眼には右方向に移動する縞を見せると、縦縞が遠ざかるように感じます。私たちはこのような刺激を痲酔下の動物に提示したときに生じる内因性シグナルの計測を行いました。
図4は、記録されたデータから先に述べた空間的にゆるやかに変化するノイズ成分を差し引いたシグナルによって構成され
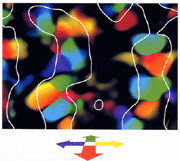
画像をクリックすると拡大します。
図4
奥行き運動方向のコラムパターン
神経活動が最も強く昂進する奥行き運動方向のドメインは色相によって表されています。白い曲線は縦縞に反応する領域の境界を表しています。
|
た奥行き運動方向のコラムパターンです。
詳細な解析によって、このようなコラム構造は単眼刺激に対する反応からは再現することができないことが確認されました。このことは、個々のニューロンの示す最適運動方向は左右眼で一致していて、ニューロン単位
では奥行き運動方向をコードすることができないことを意味しています。すなわち、奥行き方向の運動は、方位
や眼優位性と異なり、大脳皮質ニューロン間の非線形な相互作用によってはじめて抽出されると考えられます。
おわりに
現在私たちは、光計測実験や自己組織化理論で得られた知見を基に、皮質内の相互結合を仮定した大規模なニューロンネットワークのモデルを構築し、大脳皮質内の計算論の解明に取り組んでいます。このような理論と実験による相補的かつ相乗的なアプローチによってはじめて、脳の発達や視覚認知の原理が解明されるものと考えています。
|
|
|
|
|