





人間は生物の中で最も社会的な動物です。これは我々が持っている、学習によって習得した音声コミュニケーションの複雑なシステム、そして、言語によって維持される親密な個人的関係の形成において、特に際立っています。このような社会的相互関係は、精神的、肉体的健康を維持するうえで不可欠であるため、社会的行動を制御する脳のメカニズムを理解することは重要です。その動物モデルとして使われるソングバード(スズメ目鳴禽類に属する、歌をさえずる鳥)の社会的行動は、人間の社会的行動と極めて似通っています。この仲間の種の多くは、人間と同様、他個体との親密かつ終生続く友好関係を築きます。さらに、コミュニケーションのための発声を若い個体が学習する際には、その時の社会的環境が非常に大きく影響します。
当研究チームでは、歌行動における以下の2つの局面に焦点を合わせています。その1つは、限られた期間内に学習する必要がある複雑な社会的行動の一例として、歌(さえずり)の学習を研究し、そのような学習を可能にするニューラル・ネットワーク(神経回路網)および可塑性のメカニズムの特性を明らかにすることを目的としています。また、成鳥が歌のコミュニケーションを行う際に、社会的な情報が、脳の活動および鳥の行動に対してどのように強い影響を及ぼすか、という研究も行っています。
幼鳥がはるかに上手に歌を学習できるのはなぜか
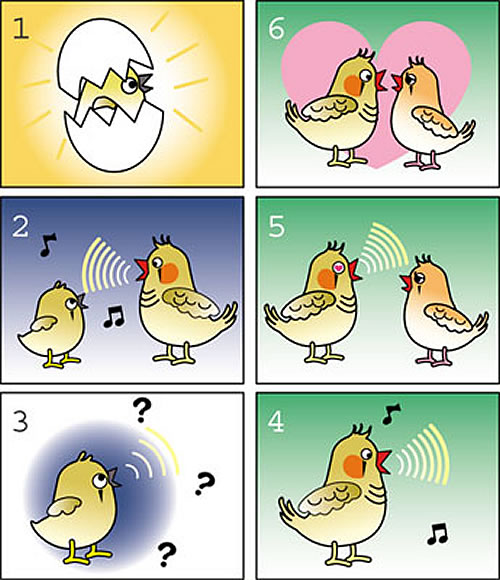
当研究チームでは、主にオーストラリア原産の小鳥であるキンカチョウを扱っています。歌を学習するために、雄キンカチョウの幼鳥は、まず父鳥が歌うのを聞き、ある程度の歌の記憶を脳内に形成します(図1)。少し経つと、幼鳥は自身で歌い始めます。練習している間、幼鳥は自身の歌を聞き、脳内にある記憶により良く符合するように、徐々に歌に磨きをかけます。このようにして、幼鳥は最終的に父鳥の歌を上手くコピーをすることができるのです。この学習は、限られた期間内に行われる必要があります。100日齢を過ぎた鳥は、もはや新たな歌を記憶したり、自身の覚えた歌を修正することはできません。このような優れた学習能力をもたらす幼鳥の脳と成鳥の脳とには、どのような違いがあるのでしょうか。幼鳥がより優れた学習機能を備えているのは、成鳥に欠けているある種の脳の可塑性のおかげでしょうか。我々は、キンカチョウの脳内で、このように年齢の限定された可塑性の探求を開始しました。
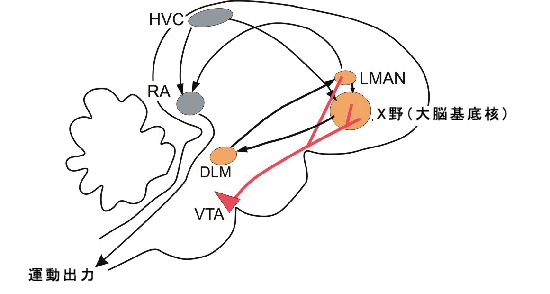
ソングバードは、さえずることのみに特化された脳内歌制御中枢を備えているため、このような研究を行う上で好都合です(図2)。つまり、歌学習に関連する重要な可塑性を解明するために、脳内のどこを調べれば良いのかが分かるので、優れた研究モデルとなるのです。そこで、キンカチョウの歌学習を研究モデルとして、脳内における特定の部位での、歌学習を制御するとされるシナプス可塑性についての研究が試みられています。興味深いことに、キンカチョウや他の多くの鳴禽類において、雄だけが歌をさえずります。そのような種では、これらの特化された脳内歌制御中枢があるのも雄のみです。我々は、これらの中枢のうち、発声中枢である神経核RAにおいて、可塑性のメカニズムを成している可能性があるものを探し始めました。この神経核内のニューロンは、歌制御中枢である神経核HVCおよび可塑性に関係する神経核LMANの両方から入力を受けています。歌学習のどの過程においても、この領域が損傷を受けた場合、鳥は歌う能力を失います。そこで我々は、歌学習を促進しうるシナプス可塑性の形を探す最初のステップとして、歌学習期における基本的なシナプス特性を明らかにすることにしました。
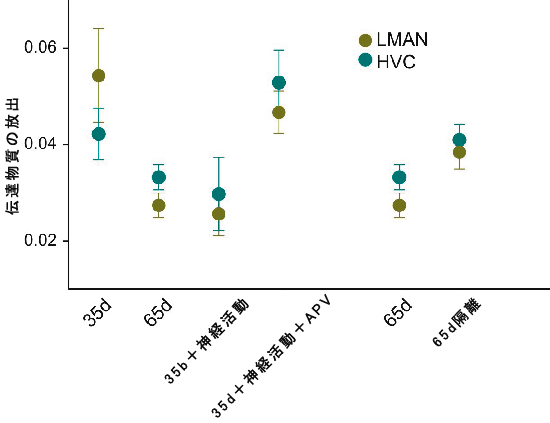
その結果、歌学習期間のRAでは、HVC・LMAN両方の神経核由来のシナプスにおいて、伝達物質の放出がおよそ50%減少することが分かりました(図 3)。このようなシナプス前機能の変化は、幼鳥からとった脳スライス内での急激な活動の増加によって再現することができ、NMDA受容体の機能をブロックすることにより阻害されました。さらに我々は、正常な歌学習を妨げられた60日齢の若鳥のシナプス前機能が、それより若い歌学習期の初期段階にある幼鳥のシナプス前機能と同様に残っていることを発見しました。このようなシナプス前機能が成熟することは、歌学習と関係している可能性を示唆しています。これらの研究の成果は、歌学習期間の幼鳥に強く見られ、歌学習をそれ以上行うことができない成鳥では減少する、という、シナプス可塑性の一つの形を示すものです。
最近、我々は、このシナプス前機能の変化が、すでに特徴がよく解明されている哺乳類の長期抑制(LTD)と同様の特性を備えている可能性があることを発見し、この可塑性が歌学習において必要不可欠なのかどうかを解明することを目指しています。
成鳥はなぜ歌うのか
若い雄鳥は、たくさんの時間と多くの脳内領域を、歌学習のために捧げます。
なぜ、歌学習がそれほど重要なのでしょうか。雄にとっては、若いうちに種特有の歌を発達させることが不可欠であり、それによって、雌鳥を引きつけて、子孫を残すチャンスを得るのです(図1)。我々は、歌うための脳の経路と、求愛に関係する一般的な動機付け回路が、どのように相互に作用しているのかを研究しています。
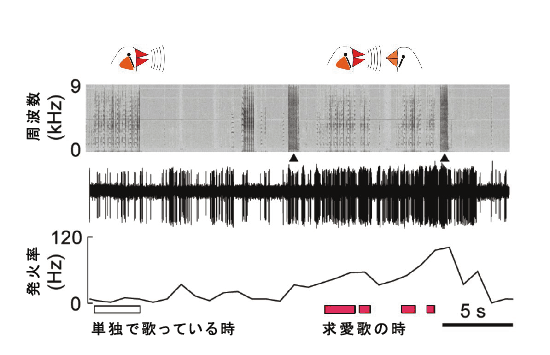
雄のキンカチョウは、単独の際も、雌に求愛をする際も、極めて似通った歌を歌います。しかし以前、我々は、いくつかの前脳領域(図2、オレンジ色の中枢)における神経活動のレベルおよびパターンが、これら2つの歌の内容によって大きく異なっていることを発見しました。つまり、この前脳における違いは、歌の「意味」に関連している可能性があるのです。この前脳の活動を修飾する可能性の一つとして、最近我々は、中脳ドーパミン系腹側被蓋野(VTA)内のニューロンが、雄が単独で歌っている時よりも、求愛のために歌う時のほうが、はるかに強く活性化されるという事実を発見しました(図4)。一般的に、雄鳥が雌鳥を目にした際、このようなニューロンの活動はわずかに変調し、雄鳥が雌鳥に対して歌う際には、さらに強く活動します。
この脳のシステムは、人間においても一般的である、報酬と動機付けに関係があると考えられています。中脳辺縁系ドーパミン経路を活性化させる報酬刺激は広範囲に及び、テレビゲームやチョコレートといったものも含まれます。おそらく、我々の研究に最も関係が深いのは、脳のイメージング実験によって、人間が自分の愛する人を目にした際に、同様のドーパミン系が活性化されるという事実が分かったことでしょう。我々の行っている神経活動の記録も、キンカチョウの脳において同様の感情の痕跡を示すことができるのでしょうか。
脳の報酬システムに関するその他の興味深い局面としては、システムの構成要素と相互に作用する化学物質によってシステムが活性化されるという点が挙げられ、これが薬物中毒の起きる仕組みであると考えられています。人間や他の動物において、アンフェタミンなど、数多くの薬物を使用することで、中脳ドーパミン系における活動が増大します。この活動は、VTAにおけるシナプスの機能の長期にわたる増大をもたらします。現在、我々は、同様のシナプス強化が、求愛歌をさえずった後の雄鳥にも起こるという予備データを得ています。
ソングバードと人間とは見た目が大きく違ってはいますが、両者の行動や社会環境には十分な共通点があり、我々の研究が、我々の社会行動の根底にある脳のメカニズムに関する知見を提供できると信じています。
本レポートに収録されているデータを提供して下さった、浅井圭子氏、王健氏、 柳原真氏に著者から感謝を捧げます。
