





Humans are among the most social of animals. This is evident especially in our complex system of learned vocal communication, and in our formation of close personal relationships maintained by language. Because such social interactions are critical in maintaining mental and physical health, it is important to understand the brain mechanisms that control social behavior. As an animal model, the social behavior of songbirds has notable similarities to that of humans. Many species, like humans, form close lifelong affiliations with other individuals. Further, the learning of communicative vocalizations by young animals is especially sensitive to a precise social setting.
Work in our lab focuses on these two aspects of singing behavior. We study song learning as an example of a complex social behavior that must be learned within a limited time period, and aim to characterize the neural networks and plasticity mechanisms that allow such learning. We also study the way in which social information strongly influences brain activity and behavior during song communication by adults.
Why are young birds so much better at learning to sing?
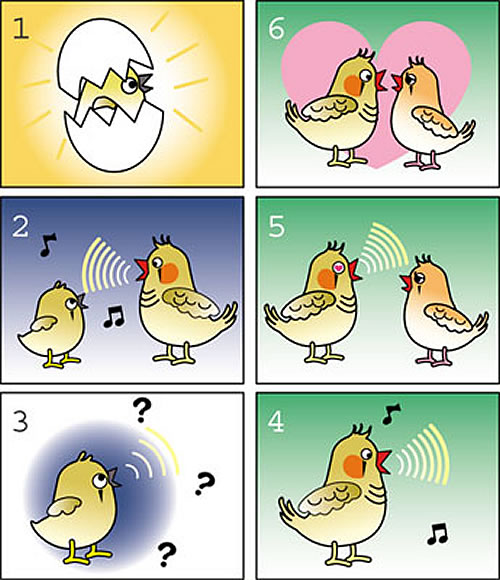
From about 25 to 50 days after birth (1), a young male zebra finch listens to his father sing, and forms some memory of this sound in his brain (2). The young bird begins to sing himself when about 30 days old, and progressively refines his initially noisy song (3) by comparing his output to the stored memory of the father's song, until he can make a good copy of the father's song by about 70 days old (4). Adult male uses this learned song to court a female finch (5), who can choose to pair with the male if she likes his song (6)
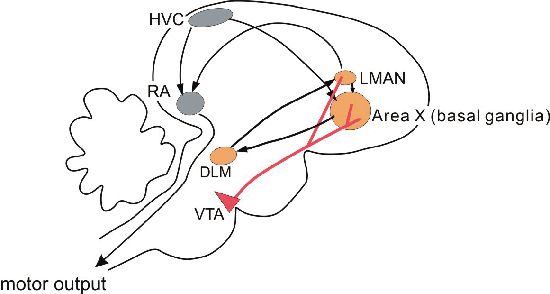
A side view of the songbird brain, with anterior to the right. Gray motor control nuclei are critical for both learning and production of song, while orange nuclei are mainly related to song plasticity. The midbrain VTA provides dopaminergic input to orange nuclei.

The probability of transmitter release at synapses from LMAN and HVC to RA decreases from early to late during the song learning period. Acute increase of activity and subsequent activation of NMDA receptors can also lead to decreased transmitter release. Preventing young birds from completing their song learning blocks some degree of maturation of presynaptic function.

The level of activity of a VTA neuron is much higher during courtship singing to a female (directed) than when singing alone (undirected). Top panel show song pattern and lower panels show neural activity trace and firing rate. Singing periods are indicated by bars beneath the bottom panel.
Our lab works mainly with the zebra finch, a small bird native to Australia. In order to learn to sing, a young male zebra finch first listens to his father sing and forms some memory of the song in his brain (Fig. 1). A little later, the young bird begins to vocalize himself. While practicing, the bird listens to his output, and progressively refines his production so that it better matches the memory in his brain. In this way, he can eventually make a good copy of the fathers song. This learning has to occur within a restricted period of time - after a bird is more than 100 days old, he can no longer memorize new songs, or change his own song. What is the difference between the brain of the young and old bird, giving the young animal such a greater capacity of learning? Perhaps young animals have a greater learning facility due to some form of brain plasticity that is lacking in adults? We have begun to search for such age-restricted plasticity in the zebra finch brain.
Songbirds are advantageous for such studies, because they have specialized brain areas dedicated only to singing (Fig 2). So, we have a good idea where in the brain we should look for critical plasticity. Interestingly, in the zebra finch and many other songbirds species, only males sing, and only males have these specialized brain areas. We began to look for candidate plasticity mechanisms in the nucleus RA. Neurons in this nucleus receive input from a singing control nucleus, HVC, and a plasticity-related nucleus, LMAN. If this area is damaged at any time, a bird loses the ability to sing. As a first step in searching for forms of synaptic plasticity that could subserve song learning, we characterized basic synaptic properties during the learning period.
We found that, during the period of song learning, the release of transmitter decreases by about 50% at synapses from both input nuclei (Fig.3). Such an alteration of presynaptic function could be mimicked by acutely increasing activity in a brain slice taken from a young bird, and this was prevented by blocking function of the NMDA receptor. Further suggesting that this maturation of presynaptic function may be related to song learning, we found that in 60 day old birds who were prevented from normal learning, presynaptic function remained similar to that of young birds early in the learning period. These findings introduce one form of synaptic plasticity that is strong in young birds during learning, and reduced in older birds who can learn no more. We have recently found that this alteration of presynaptic function may have similar properties to well characterized forms of long term depression (LTD) in mammals, and are further testing directly whether this plasticity is critical for song learning.
Why do adult birds sing?
Much of the young male songbird's time, and a large portion of his brain, is dedicated to learning to sing. Why is this so important to him?it is critical that the male develops a proper song, so that he can attract a female finch and have a chance to produce offspring (Fig. 1). We are investigating how the brain pathways for singing interact with general motivational circuits related to courtship.
Zebra finch males sing a very similar song while alone and during courtship with a female. We previously found that the level and pattern of neural activity in several forebrain areas (Fig. 2, orange nuclei) are very different during these two singing contexts. This brain difference could thus be related to the "meaning" of the song. As a possible source of this forebrain modulation, we recently found that neurons in the midbrain dopaminergic ventral tegmental area (VTA) are much more strongly activated during courtship singing than when males sing while alone (Fig 4). Typically, activity of such neurons was slightly modulated when males saw a female bird, and further more strongly modulated when the male also sang to the female.
This brain system is thought to be involved in general reward and motivation in humans, as well. A wide range of rewarding stimuli can activate the mesolimbic dopamine system, including things such as video games and chocolate. Perhaps most relevant for our studies, brain imaging experiments have found that the same systems are activated when humans see someone they love. Could our recordings represent traces of a similar emotion in the zebra finch brain?
Another interesting aspect of the brain reward system is that it can be activated by chemicals that interact with components of the system. This is thought to be the means by which drug addiction occurs. In humans and other animals, the use of many drugs such as amphetamine causes increased activity in the midbrain dopamine system. This activity leads to a prolonged enhancement of function of synapses in VTA. We now have preliminary evidence that a similar synaptic enhancement occurs in male songbirds after courtship singing.
Although songbirds may seem very different from humans, we believe that their behavior and social environment has enough in common with ours that our research may provide some insight into the brain mechanisms underlying our social behavior.
The author thanks Keiko Asai, Jian Wang, and Shin Yanagihara for providing material included in this report.
