





21世紀に入り、ヒト、マウスを含むいくつかの動物種において、その全ゲノム領域の配列(一部確定していない部分もある)が決定されてきました。それに伴い、たくさんの遺伝子が同定され、さらに、産物としての蛋白質をつくることのない、多くのRNAが染色体上にコードされていることが見出されてきました。
一つの染色体というのは100万単位の膨大な核酸の集合体であり、これら数多くの遺伝子(あるいはRNA)はDNA巨大分子である染色体上でどのような機構で発現(=転写)の調節を受けているのか、というのが我々の研究のテーマです。
免疫系の細胞など一部の細胞を除いて、ある生物の一個体において全ての細胞は同じ染色体を持っていると考えられています。しかしながら、それぞれの細胞で常に全ての遺伝子が発現しているという訳ではなく、必要に応じて、遺伝子は染色体から読み込まれて発現し、機能を発揮します。
例えば、胎児期の神経系ではその時期特有の、大人の脳ではまたその時期の脳特有の遺伝子群がそれぞれの細胞で発現しています。一方で、その時期、その細胞で発現しているべきではない遺伝子が発現することで、形態の異常や病気などのさまざまな障害が起こる場合もあります。それを防ぐためにも、個々の遺伝子は正確に読み込まれる必要があります。
遺伝子の発現調節の機構として一般的に受け入れられているのが、転写の開始点を規定するプロモーターと呼ばれているDNAの配列(領域)と、そのON-OFFを実際に決定する領域であるエンハンサー、あるいはシス転写調節領域と呼ばれるDNA配列(領域)の組み合わせにより、特定の細胞での遺伝子の発現(転写)が決定されているという説です。
しかしながら、染色体という巨大なDNA分子の中には、たくさんの遺伝子、プロモーター、そしてエンハンサーとして働きうるDNAの配列が存在すると考えられ、その中でそのプロモーターおよびエンハンサーという二つのDNA領域がどのように相互作用するのかに関しては全く分かっていません。
近年の遺伝学の進展により、遺伝子の調節領域が数百キロベースあるいは1メガベースという膨大な距離に離れて存在する場合も知られて来ています。そのような染色体という環境の中で、転写調節領域がプロモーターに作用するために必要とされる機構や、転写調節領域がその調節下に置くことができる範囲の解明、すなわちその範囲を決定する機構に関しては、現在ほとんど知られていません。
発生過程においては細胞の増殖、分化が非常に速い速度でダイナミックに進んで行きます。その中でたくさんの遺伝子が発生における生育時間特異的に、そして組織特異的に発現し、働きます。したがって、それらの遺伝子は発生の過程においてダイナミックに発現のON-OFFを行うことになります。そのON-OFFを間違えれば、組織の形態、ひいては機能の異常を引き起こします。
染色体という巨大なDNA分子の中で、間違えることなくその調節を行うための機構とはどのようなものなのか、我々は身体の前後軸における特異性を決定する因子であるHox遺伝子群を主な研究の対象として用いてその調節の機構の解明を目指しています。
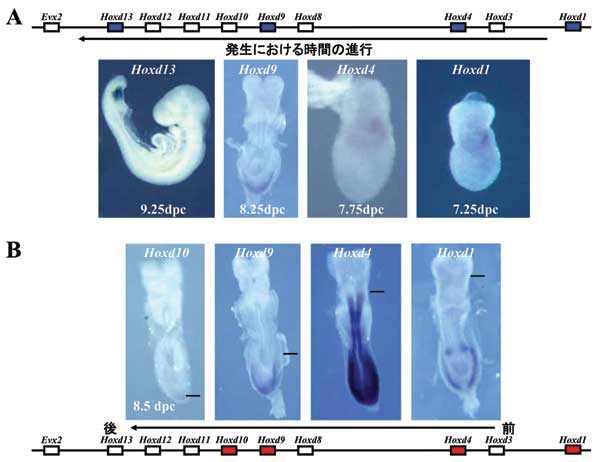
A: 発生の時間におけるHoxd1、4、9、13の発現開始(青で示してある遺伝子)。遺伝子の発現の開始時期は染色体上に並んでいる順番に対応する。
B: マウス受精後8日胚でのHoxd1、4、9、10の発現(赤で示してある遺伝子)。発現領域の最前部に線を引いてある。この時点でHoxd11からEvx2は発現していない。
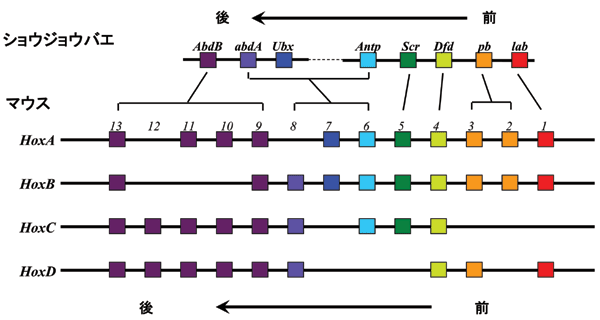
Hox遺伝子はショウジョウバエの遺伝学の成果として、動物の形態形成に重要な役割を果たしている遺伝子群の最初の例として発見されました。ショウジョウバエでは8個、ヒト、マウスなどの哺乳動物では39個の遺伝子が見つかっています(図1)。
それらは全てホメオボックスと呼ばれるDNA結合ドメインを持つ転写調節因子であることが分かっています。これらの遺伝子は染色体のある部分に集合して存在しているという特徴があります。哺乳動物の場合では約10個ずつの遺伝子が同じ転写方向に、約100kbの長さの4カ所の染色体領域に集合して(クラスター状に)存在しています(コンプレックスと呼ばれている)(図1)。
この遺伝子群はそれぞれのコンプレックスにおいて、遺伝子の並んでいる順番が遺伝子の発現の順番を反映しているという面白い特徴を持っています。一方の端(3'と呼ばれています)の遺伝子がより発生上の早い時点で(マウスでは受精後7.0日齢位から)発現が始まり、染色体上の順番を追って行く形で順番に遺伝子が発現して行き、最後にもう一方の端にある遺伝子が発現する(マウスでは受精後9.5日齢位)、という形になります(図2)。
また、同様に、遺伝子の発現領域においても遺伝子発現パターンは染色体構造を反映する形になっています(図2)。これらのことから、Hox遺伝子群は非常に密集して存在しているにも関わらず、各プロモーターに作用するエンハンサーは厳密に選択されていることが分かります。また、Hox遺伝子コンプレックスの中ではなく、300kbもの距離にエンハンサーが離れて存在している例も見出されています。
遺伝子の染色体上での存在位置と発現調節の関係については、さまざまな努力により明らかにされつつあり、時間的な調節はコンプレックス全体を一括して調節する機構に主として依存していることが解明され始めています。また、エンハンサーとプロモーターの相互作用に関するルールも徐々に明らかになりつつあります。しかしながら、未だにそれだけでは説明のできない矛盾も多々含んでおり、今後さらにそれらを一つ一つ解きほぐして行かなくてはなりません。
