




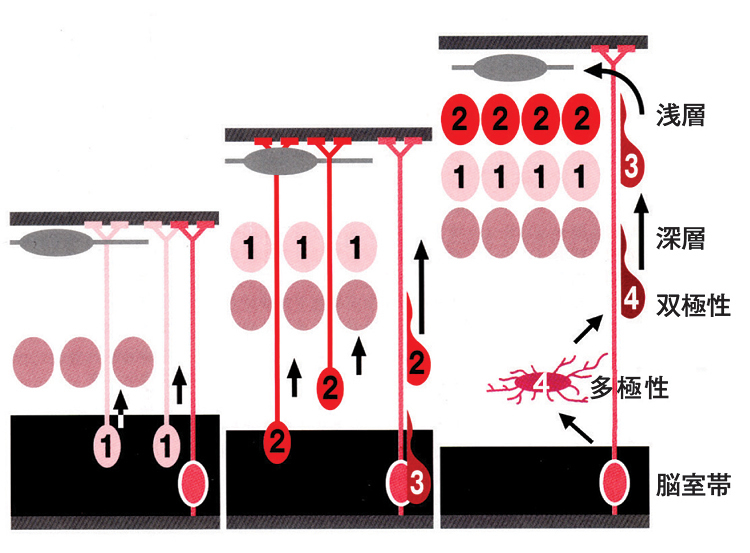
大脳皮質を構成する錐体細胞は、胎生期に大脳の内側の脳室帯に多数存在する神経前駆細胞が分裂し、神経細胞となったもので、表層に向かって放射状に移動する。錐体細胞の放射状移動は、早く生まれた細胞が深層に、後で生まれた細胞が浅い層に位置する「インサイド・アウトパターン」を取りながら、皮質を形成する。数字は生まれ順を示す。移動開始の早期に、多数の突起を伸縮する「多極性」と呼ばれるステージがあり、その後に先導突起と将来の軸索に相当する2つの突起のみを有する「双極性」という形態に変化する。双極性の形態を獲得した後は、放射状グリアと呼ばれる構造に沿うように移動する。
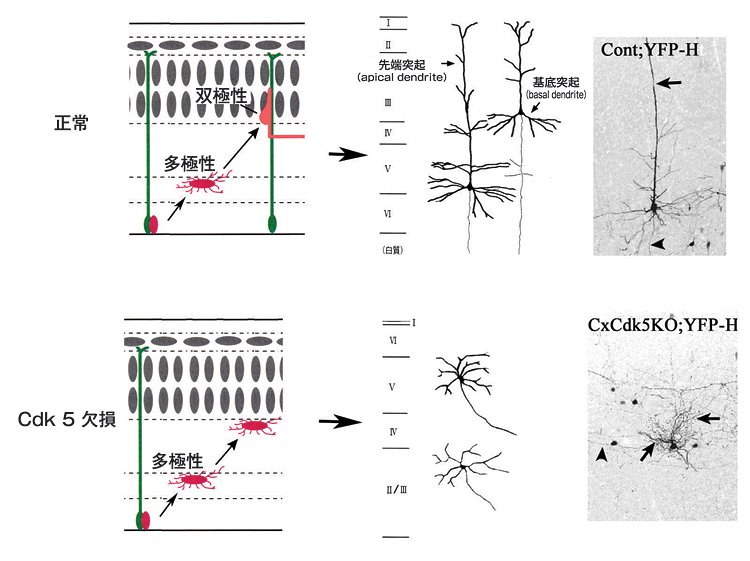
Cdk5欠損マウスでは、図1に示したインサイド・アウトパターンの皮質形成が起こらず、II-VI層の位置が逆転している。変異マウスの異常から、正常な大脳皮質形成では、神経細胞が多極性から双極性に形態を変化させる過程そのものが、脳でみられる特徴的な神経細胞の形づくりの基盤になっていることがわかった。
私たちの脳は、極めて精緻で、整然とした構造を持ち、驚くべき能力を発揮します。脳が正常に機能するためには、正しく形づくられる必要があります。哺乳類の大脳皮質は、規則正しい6層の構造をしています。この6層の構造は、神経細胞が増殖する場所から、最終的に配置される場所へ自ら移動し、配置されるという巧妙なメカニズムにより形成されます。その際、早くできた神経細胞がより深層に、後からできた神経細胞が浅い層に分布する「インサイド・アウトパターン」を取りながら、皮質を形成します。移動する際、神経細胞は、多数の突起を有する多極性という形態から、2本の突起を有する双極性に変化することが最近の研究からわかってきました(図1)。また、大脳皮質を形成する「錐体型神経細胞」は、先端突起と呼ばれる1本の長い樹状突起を脳の表面に向けて伸ばすことが知られています。しかしながら、こうした形態が、どのようにして獲得されるかは、まだ良く判っていません。
我々の研究グループは、神経細胞の移動や細胞の形態変化のメカニズムを探るために、細胞内のタンパク質をリン酸化する酵素サイクリン依存性キナーゼ5(Cyclin-dependent kinase5、Cdk5)に着目しました。Cdk5欠損マウスにおいて大脳の構造が異常になることを発見していたので、大脳皮質の形成時に、神経細胞が形態を変化させる様子を、正常なマウスと、Cdk5を大脳皮質で働かなくしたマウスとで詳細に検討しました。その結果、Cdk5を働かなくしたマウスでは、多極性から双極性への形態変化を起こさないとともに、神経細胞が生まれた場所からあまり移動しないことがわかりました。さらに錐体型神経細胞は、先端突起を脳の表面に伸ばしませんでした。これらの結果から、Cdk5が、この大脳皮質における神経細胞の形態変換に重要であり、形態変化が起こらないと、神経細胞の移動が効率良く起きないことがわかりました。このことから、正常での形態変換により、錐体細胞の形態が形づくられる事が推定されました(図2)。Cdk5のリン酸化が必要なターゲット蛋白質の侯補はいくつかありますが、今後神経細胞の移動過程での形態変換に直接関与する蛋白質を同定する事で、さらに詳しいメカニズムが明らかになる事が期待されます。
