






われわれの知覚系は正常な状況のもとでは、外界の情報を正確に提供してくれる。しかし、視覚がわれわれを誤った結論に導くか、曖昧な結果をもたらすような局面も数多く存在する。「多義図形」は視覚系の心理学的研究で最も広く知られた図形のひとつである。例えば、アトニーブの三角形(図1)を眺めていると、最初のうちはすべての三角形がある特定の方向に向かっているようにみえるが、しばらくすると、別の方向を向いているようにも見えてくる。そしてこのような現象(知覚交替)が無限に繰り返される。
例えば、三角形が向きを変えるときは、すべての三角形が一斉に向きを変えるように、このような知覚交替(多義図形の「見え」が変化する現象)は「全か無か」の現象であると考えられてきた。しかしアトニーブも指摘しているように、このような現象がいつもみられるとは限らない。ときには三角形は方向性を失ったり、それぞれが思い思いの方角を向いたりして、全体像の印象を弱めてしまうことがある。証明することは困難であるが、同様な現象が、例えばよく知られているネッカーキューブのような多義図形にも起こることがある。視覚系は全面的整合性をわれわれが期待するほど厳密に維持するわけではない。
悉無的に、あるいは部分的にかにせよ、図形の「見え」が変化することは交替現象の発現において重要な役割を演ずる。事実このことは、交替現象を説明する数多くの神経理論では前提とされていた。ゲシュタルト心理学者ヴォルフガング・ケーラー(Wolfgang Kohler)は、この図形の「見え」の変化は神経系の疲労によるものであると考えた。現在では、神経系の相互に同期した活動が不安定性を有することが、パターン変化の原因であると考えられる場合が多い。これらすべての認識は、一元的で、低次の(脳内)情報処理機構が交替現象の根底にあるという前提に基づいている。しかしながら、われわれ研究チームが最近行った研究では、知覚交替に関連した情報処理が順に生じることにより、図形の「見え」が変化するということが明らかになった。
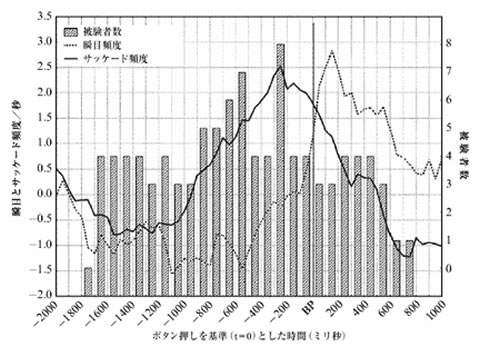
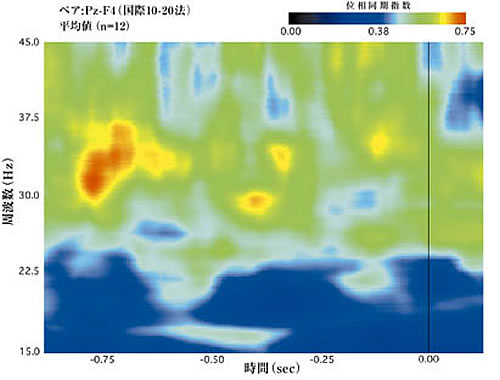
図2は、知覚交替に先行して現われる一連のイベントを、眼球運動と脳波それぞれについて示したものである。図2Aでは被験者は図のすべての三角形の向きが変わるのに気付くたびにボタンを押す(BP)ように指示された。瞬目(目を閉じ、またすぐに開くこと)とサッケード(一方の注視点から他方の注視点への衝動的な眼球運動)の頻度、および三角形の見かけの向きに整合性がある眼球運動が記録され、サッケード運動の方向と、図1の三角形の見かけの向きが一致する頻度がそうでない場合に比べて有意に大きいことがわかった。交替現象の発生の1700ms~700ms前では瞬目とサッケードは抑制されている。文献によればこのような現象は認知活動と関わりのある現象として理解されている。したがって、認知活動が交替現象を開始すると推論してもよいであろう。この時期が過ぎ、ボタン押しのおおよそ700ms前から、次に知覚される三角形の向きに一致する方向の眼球運動の頻度の上昇が認められるようになる。このような眼球運動は新しい向きに対する自覚的意識と関連している可能性がある。ボタン押しの後の瞬目頻度は通常より多く、これは1700ms~700ms前に抑制されていたことに対する補償の一形態と思われる。
図2Bは、ネッカーキューブ(Necker cube)を見ているときの知覚交替に対応して現れた同期した脳波活動を時間周波数領域上でグラフ化したものである。暖色濃度が高いほど、頭頂部と前頭部の間の同期性が高いことを示している。このプロットでは高いコヒーレント性によって特徴付けられる3つの短い神経活動がみられる。第1は、ボタン押しの 700ms前付近で、図2Aのサッケードの頻度が再び上昇を開始する時間に対応し、眼球運動についての我々の解釈と一致する。これは次の「見え」が確立されつつあることを示している。2番目の同期した活動は、ボタン押しの350ms前で、三角形の見かけの向きと方向が一致した眼球運動の頻度が最大になる時刻に一致する。したがって、これは次の「見え」によって注意が捕捉されたことを表わしているといえよう。最後に現われる同期した神経活動はボタン押しの 120ms前に起こる。これはボタン押しに伴う運動性の機能に対応する。
図2では、知覚交替が認知イベントを伴い、感覚イベントが続き、運動イベントがさらに続くことを示している。イベント(神経活動)のこのような一連の流れはトップダウンの形であらゆる皮質機能を巻き込んでゆくことを示している。これは、われわれの知覚交替を意識的にコントロールすることが可能か否かという興味ある問題を提起する。知覚交替しないよう努めることを被験者が指示されると、知覚交替の回数はさらに減少する(知覚交替の発現を知らせることを単に躊躇している状態と区別するよう注意する必要があるが)。多義図形のいくつかの解釈(見え方)の中でどれか1つの解釈を保持する(hold)ように指示されると、ある限られた時間は指示された通りに行うことができる。しかしこのことによって被験者がもう一方の解釈に知覚交替することが妨げられることはない。コントロールは従って限られた範囲でのみ可能である。
知覚交替は、視覚系は能動的なシステムであるという重要な例証をわれわれに提供してくれる。知覚系は必ずしも新しい情報が入力されて初めて次の状態に変化するとは限らない。ときには自分でその状態を変えてしまうこともある。ここで示された結果は、知覚交替はより高次の視覚機能から始まり、低次の視覚野内での神経活動の不安定性によるものではないことを示唆している。
