





抑制性ニューロンの研究は、興奮性ニューロンの研究とならんで必要です。われわれの目標は、抑制性ニューロンの神経伝達物質であるGABA(ガンマ・アミノ酪酸)について全体像を明らかにすることです。脳のニューロンのうち約30パーセントはGABA性の抑制性ニューロンで、このニューロンだけがGABA合成酵素のグルタミン酸デカルボキシラーゼ(GAD)を持っています。このほかに小胞性GABAトランスポータ(VGAT)、細胞膜性GABAトランスポータ、GABA分解酵素をセットにして持っており、シナプス前神経終末でGABAを合成して、シナプス小胞に詰め込み、細胞外(シナプス間隙)で用済みになったGABAを取り除き、余分のGABA を分解してGABAの量をコントロールしています。
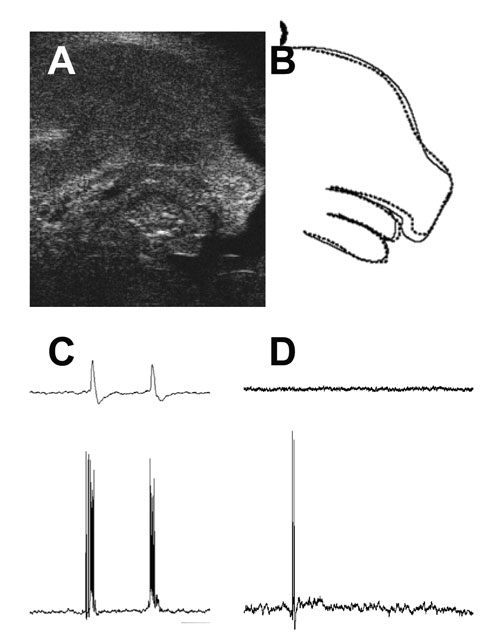
AとB:子宮内マウス胎児(14日目)の超音波画像(A)と模式図(B)。点線は口が開いたとき。正常マウスはときどき口を開けるが、GAD67ノックアウトマウスはほとんど開けない。
CとD:マウス胎児(18日目)の脳幹で記録した呼吸性電気活動。正常マウスでは(C)、ニューロンが呼吸リズムで発火し(下のトレース)、同期して運動神経が活動する(上)。GAD65と67の両方をノックアウトすると(D)、ニューロンはときどき発火しているが(下)、運動神経は活動していない(上)。
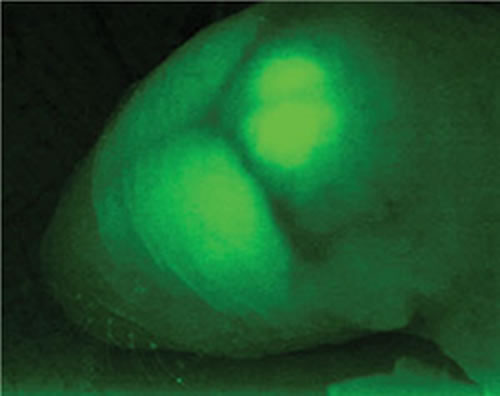
われわれは2種のGAD(GAD65と GAD67)とVGATの遺伝子ノックアウトマウスをつくり、BSI内外の研究室と共同研究しながら、GABAを減らして起こる異常を見ることで、 GABAが持つ役割を調べています。また、蛍光蛋白質GFPの遺伝子をGAD67遺伝子にノックインして、生きたままのGABAニューロンをGFP蛍光で見分けられるマウスもつくって利用しています(図1)
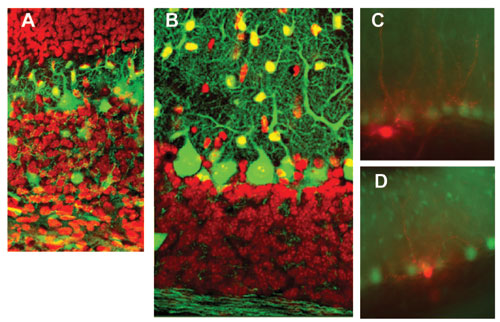
大脳皮質ができるとき、興奮性ニューロンと GABA性の抑制性ニューロンとが別々のところで産まれ、別々の様式で移動してきます。われわれは、GFPで光るGABAニューロンが移動して行くのを連続撮影して詳しく解析しました(大阪大学村上研究室との共同研究)。小脳皮質では、興奮性の顆粒細胞が表面でつくられてから深部へ移動し、残りのプルキンエ細胞と抑制性介在ニューロン(すべてGABA性)は脳室側から表面へ移動して、小脳特有な構造ができます。われわれはGFPマウスで、プルキンエ細胞と顆粒細胞の発達に合わせて、介在ニューロンが白質で増殖しながら皮質に送り込まれるのを示しました(図2A、B)。GFPマウスを使った電気生理学実験で、GABAニューロンが視床下部のホルモン分泌調節(米国エール大学との共同研究)や、脳室周囲器官での体内ナトリウム感知(基礎生物学研究所との共同研究)などにはたらいていることを示し、小脳皮質でGABA性神経伝達がアドレナリンなどによって調節されるメカニズムを調べています。数が少なく、研究が進んでいない介在ニューロンのルガロ細胞、カンデラブルム細胞についても調べています(図2C、D)。
GAD65ノックアウトマウスでは脳の GABAが半分に減って、てんかん発作が起こり、不安や恐怖などの情動が異常です。胎児期にはGAD65が未発達で、GABAは主にGAD67で合成されているので、GAD67ノックアウトマウスは誕生時にGABAが10パーセント以下に減って、神経活動も異常です。胎児でも自発的に呼吸活動や四肢の運動をしていますが、ノックアウトマウスではほとんど見られません(図3)。このマウスは、口蓋裂のために生まれるとすぐ死ぬので、生後の異常は調べられないのが残念ですが、口蓋がつくられるのを補助する舌の運動がないため、口蓋裂になると考えています。
GAD65ノックアウトマウスと、GAD67ノックアウトマウスを交配して、両方のGADがないマウスをつくり、VGATのノックアウトマウスもつくりました。これらのマウスではGABAの作用がすべて失われ、脳幹や脊髄で運動神経のリズム活動がまったくありません(図3D)。二次的に運動ニューロンの数の調節が妨げられて、余分のニューロンが残ります。GABAのはたらきで見つかっていない現象や分からないメカニズムはまだまだ多く、研究に終わりはありません。
GABAは動物では脳に特有のアミノ酸として発見されましたが、以前からGADとGABAを植物や微生物が持つことは知られていました。最近、植物で雌しべで起こる受精および紫外線障害の防止に、GABAが重要なはたらきをすることが分かりました。動物でもほかの一部の細胞(すい臓、毛根、生殖器など)でGABAが見つかって、今後、そこでのはたらきも分かってくるでしょう。同じ物質でもさまざまな局面で、違った役目を担当するのが生物の巧みなところであり、GABAもその一例です。必要なものはその場所でつくって使うのも生物の原則で、余分のGABAを摂取しても脳への効果は期待できません。GABA を含んだ製品が宣伝、販売されていますが、GABAはジャガイモやトマトなどに含まれていて、食物からかなり摂取されています。腸で吸収されても肝臓などでただちに分解されるので、脳に行ってはたらくことはないでしょう。

