





Introduction
Assemblies of nerve cells in the brain facilitate learning and memory. Although synaptic connection among nerve cells are widely thought to contribute to functional selectivity of these nerve assemblies, it is uncertain whether this is enough to understand the programming in the brain. Rhythmic neural activity is another collective property very frequently observed in the brain. It is an open question whether rhythmic activities can contribute to programming of the brain. Our lab examined the role of theta rhythm in the hippocampus, theoretically and experimentally, to better understand how the instantaneous property of rhythmic activity benefits the brain.

Theta rhythm in the rat hippocampus
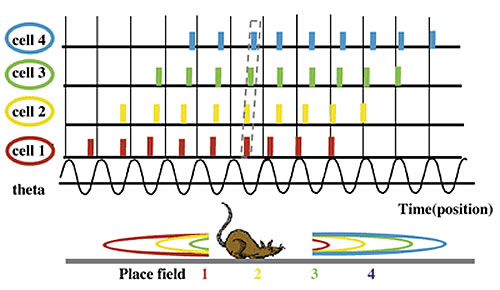
In freely-running rats, a regular, stable theta oscillation is observed in the hippocampus between 4-12 Hz of the local field potential (LFP). The predominant view of spatial representation in the rat hippocampus is the cognitive map theory (O'Keefe and Nadel, 1978). According to this theory, any given set of cells that act selectively according to the rat's place field represents an external map. These cell sets are called cell assemblies, and each individual cell within an assembly that fires at a specific location that corresponds to a unique point in the environment is called a place cell. O'Keefe and Recce (1993) first defined the relationship between the place cells and the theta rhythm as the firing pattern of individual place cells that occurs while an animal moves through an environment-or theta phase precession (Fig. 1). Skaggs et al. (1996) showed that this phenomenon is a property of place cell assemblies. Place cells activate in sequence according to the position and movement of the rat. Initial firing of a single place cell occurs at a specific phase in the LFP. Every time the cycle advances, the firing phase shifts forward. Cells that start activity near the beginning of a phase complete their cycles before those cells that start later. This maintains phase precession during activity. Throughout a theta rhythm cycle, the firing order of place cells is consistent with the running direction around the animal's current position. As a result, the duration of the temporal sequence is compressed ten-fold and embedded in the cycle of the theta rhythm.
Theta phase precession creates memory nerve circuitry
Relevance to learning initially attracted researchers' interest in phase precession in the rat hippocampus. According to our observations, the basic properties of the rhythm interaction system, known as the entrainment phenomenon, appear to govern this phase precession. If this is so, it is possible that the rhythmic pattern appears in the absence of memory to cause memory formation. We proposed a model for the dynamics of the hippocampus in which synaptic plasticity is produced selectively within the hippocampus in a way in that mirrors the temporal pattern by which cell assemblies are formed-via the interaction between the nerve cell rhythm and the LFP theta rhythm-thus allowing the pattern to be sent to the hippocampal nerve circuit. Synaptic plasticity in hippocampal pyramidal cells is not entirely Hebbian because synaptic strength increases when the presynapse and postsynapse fire with a time variance of several tens of milliseconds. Selective and unilateral synaptic formation results when hippocampal cells are activated by a phase precession that enables plasticity. The dominant hypothesis suggests that cells at the input region of the hippocampus are activated during cycle of the theta rhythm and that the range of the cycle expands gradually with the continuation of activity as more cells are voluntarily recruited. Nerve cells have been reported to become active in the cycle around the theta rhythm in the entorhinal cortex-the entrance to the hippocampus-and in the perirhinal regions. These cells have also exhibited changes in their oscillation frequency, while more thorough investigations are required.
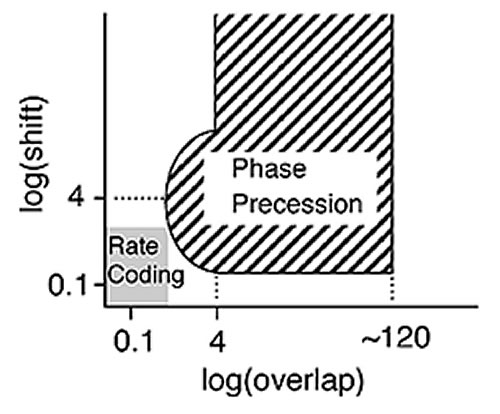
Quantitative comparison using computer experiments showed a distinct difference in the storage abilities for a correct time series when phase precession or stationary firing was involved (Fig. 3). This precession model bridges the gap between the fixed value of time interval (several tens of milliseconds) that is recognized by the synapse and the experienced event (seconds). A variety of input time series are translated by the theta rhythm, thereby giving the synapse the proper temporal difference and enabling experienced events to be encoded on the spot.
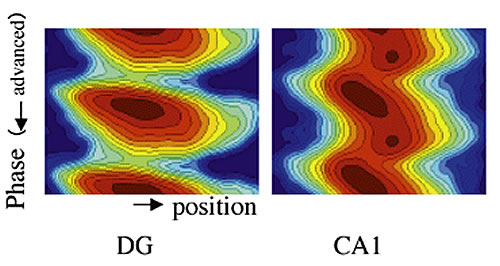
Based on our hypothesis, we conducted more analysis of phase precession interactions in subfields of the hippocampus (Fig. 4). An examination of the uniform phase shift component clearly showed a simple phase shift regulating the activity of the dentate gyrus near the hippocampal entrance that advanced in phase by a 0.2 theta cycle relative to CA1. This corresponded with the behavior predicted by the model thus showing that phase precession satisfies the conditions for memory generation. Spatial cognition in rats occurs in the hippocampus. The nerve circuit, which memorizes an environmental arrangement as a cognitive map, is produced through the natural course of time series integration. Furthermore, comparisons with other electrophysiological findings suggest that the activity of hippocampal cells other than the uniform phase shift (seen at CA1, Fig. 4) is relatively independent of memory formation, suggesting another function (e.g. coupling with concurrent executive properties).
Toward understanding the dynamics of the global brain network
The hippocampus in humans is thought to be involved in episodic memory. The above study of hippocampal theta will provide a possible neural correlate of memory formation in the episodic memory. On the other hand, brain-imaging studies showed that both the hippocampus and frontal areas are concerned with episodic memory. In such processing, the global network must be controlled for strategic or goal-oriented memory processing. Is the theta rhythm restricted to hippocampal neural dynamics or does it emerge in the global network? Interestingly, it was reported that rat hippocampal theta synchronizes with neural activities in several areas. Human theta rhythm was first observed in frontal midline area by scalp electroencephalographic measurement during mental tasks. Theta rhythm in the whole brain would provide a fascinating key to interpreting dynamical programming in the brain.
