





はじめに
記憶、学習といった機能は脳の神経細胞が集団としてもたらす働きであり、神経集団のプログラムを与えるのは、シナプス結合によってできる神経回路の選択的な構造にあると広く考えられている。一方、脳波などの測定に見られるように神経活動は絶えずリズム活動を示している。リズムという時間的な振舞の集合現象は、脳のプログラム作りに新たな機能を付加するのだろうか?以下では、海馬におけるリズム活動の役割について実験と理論の両面から、その役割を探る。

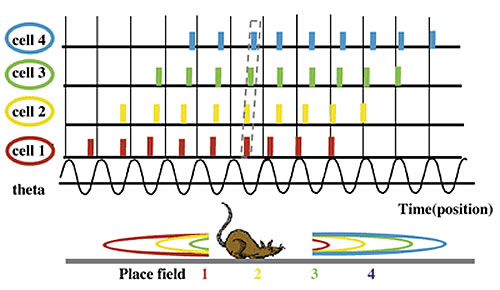
ラット海馬シータリズムの出現
自発行動時のラットでは、海馬のシータリズムは細胞外電極によりlocal field potential(LFP)の4~12ヘルツの比較的安定なリズム活動として観察される。ラット海馬の主要な働きは、空間認知地図説(O'Keefe and Nadel, 1978)として知られる。ラットの居場所に選択的に細胞が活動し、細胞集団として外界の地図を表現するとする説で、そのような細胞は場所細胞と呼ばれる。場所細胞とシータリズムの興味深い関係はO'Keefe and Recce (1993)により報告された。この最初の報告は、個々の場所細胞の発火に関するものであったが、Skaggs et al.(1996)により場所細胞集団の刻むリズムの性質として呈示され、シータ位相歳差と呼ばれた。図1に模式的に示す。ラットが走るとラットの位置の移動に応じて、場所細胞が次々活性化する。個々の場所細胞の発火を見るとLFPの特定の位相で開始し、周期が進む毎に、前にシフトした位相で発火する。早く(より出発点に近い位置で)活動を始めた細胞は後に(より離れた位置で)活動する細胞の位相を追い越すことはなく、活動中一定の位相差が保たれる。シータリズムの各1周期の間には、活性化した場所細胞が、位置の順番に従った位相の順番で発火する。その結果、ラットの「走る」という行動の時系列は、時間スケールで10倍程度圧縮してシータリズムの周期の中に埋め込まれる。
シータ位相歳差がもたらす記憶の神経回路
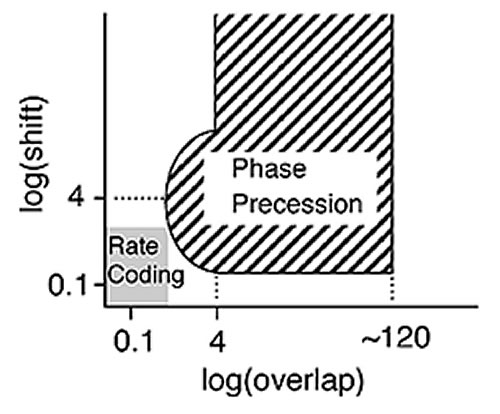
ラット海馬のシータ位相歳差は学習との関連が当初から注目された。空間を学習した回路があれば、位相歳差ができるというしくみは提案されたものの、これでは、記憶形成の役にたたない。私達はこの現象が引込現象として知られるリズム相互作用系の基本的な性質から構成できることに注目した。神経細胞のリズムとLFPシータリズムとの相互作用によって、細胞集団としての時間パターンが形成され、それが海馬の神経回路に伝えられることで、海馬内のシナプス可塑性を選択的に生みだすような海馬のダイナミクスのモデルを提案した(図2)。海馬錐体細胞のシナプス可塑性は、純粋なHebb 型ではなく、前シナプス細胞、後シナプス細胞が数十ミリ秒の時間差で活動した時に選択的なシナプス増強が起こる。位相差に従って活動した海馬内細胞同士が、可塑性に適した時間差を持つとき、選択的で一方向的なシナプスが形成されるのである。位相歳差生成の鍵となる仮定は、海馬への入力部位にある細胞が、シータリズム程度の振動数で活性化し、その自発振動数が、活動持続にともなって徐々に増加することである。海馬の入口である内嗅野およびその周辺では、シータリズム付近の周波数で活動し、しかも周波数が変化する神経細胞がすでに報告されているが(Egolov et al. 2002など)、厳密な検証については今後に待ちたい。
計算機実験による定量的な比較から、位相歳差のある場合と定常発火になる場合と比べると、図3に示すように時系列の正しい貯蔵能力に明らかな差がでることがわかった(図3)。記憶装置としてのこのモデルの利点は、シナプスの識別する固定した時間差(数十ミリ秒)と、経験される出来事(~秒)のギャップをつなぐことにある。多様な入力時系列は、シナプス固有の時間差へとシータリズム上で変換され、始めての出来事の経験がその場で記憶貯蔵されるのを可能にしている。
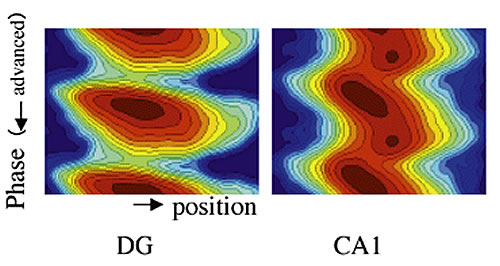
さらに、本モデルから推定される海馬の部分野に現れる位相歳差の相互関係について、実験データを解析しなおして比較した(図4)。一様な位相シフトの成分に注目すると、海馬の入口に近い歯状回の活動は単調な位相シフトのみで構成され、CA1の位相に比べて0.2周期分先行している。このことはモデルの挙動と一致しており位相歳差が記憶生成の原因として働く条件を満たしていることがわかった。
さて、ラットではもともと知られた海馬の働きとされる空間認識であった。上記のような神経回路では、認知地図としての環境の配置を記憶する回路が時系列の統合として自然に生まれる。さらに様々な電気生理で知られていた様々な知見が、理論的記憶生成のために必要な性質として比較検討できるようにもなった。これらの結果は、ラット海馬のシータリズム依存的な活動が、様々な出来事を時間空間構造として、海馬に一時的に蓄える装置を提供していることを強く支持するものである。
脳のグローバルなネットワークの理解に向けて
ヒトでは海馬はエピソード記憶に関与するとされている。ラット海馬で得られた記憶の神経機構が働いている可能性は、その記憶貯蔵能力の必然性から示唆される。しかし、ここでは海馬内に限定したモデル化であり、海馬以外のダイナミクスとあわせたよりグローバルなネットワークとしての解明が今後の課題である。ラットでも海馬だけでなく、辺縁系のいくつかの部位で同時にシータリズム活動が生ずることが知られている。ヒト脳波測定による知見では前頭中心部のシータリズム(Fmシータ)が知られており、認知思考課題中に発生することから、シータネットワークによる記憶、思考をあわせたネットワークが形成されるかどうかは、今後の重要な研究課題である。脳のプログラム作りのしなやかな性質が今後実験と理論の有効な協力により進むことが期待される。
