




背景
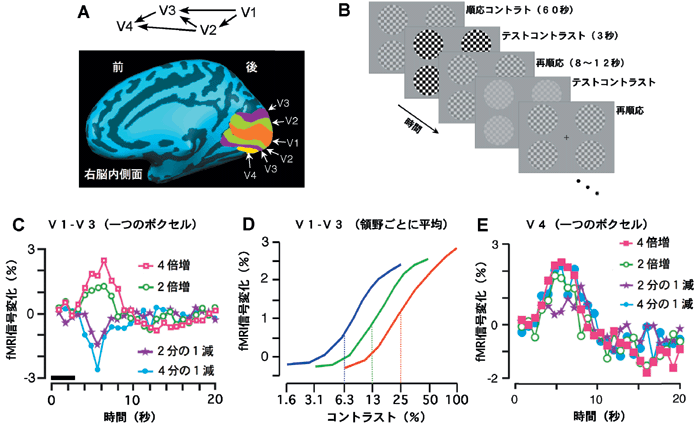
図B:コントラスト変化に対する応答をいろいろな順応状態で調べる実験パラダイム。
図C:V1野、V2野、V3野に含まれるボクセルで記録されたコントラスト変化に対する応答の例。
図D:応答のピーク値を順応刺激を提示する前の信号レベルを基準にプロットしたコントラストー応答関数。異なる色の線で3個の異なる順応状態からの応答を示す。
図E:V4野に含まれるボクセルで記録されたコントラスト変化に対する応答の例。
霊長類の大脳皮質では、網膜からの視覚信号は後頭葉の一番後ろにある第一次視覚野(V1野)で処理され、さらに高次の視覚野[第二次視覚野(V2野)、第三次視覚野(V3野)、第四次視覚野(V4 野)]で処理が加えられた後(図A)、下側頭葉や頭頂葉の視覚連合野へ送られます。
サル視覚野の神経細胞活動を記録しながらいろいろな明暗対比(コントラスト)の刺激を与えると、弱いコントラストでは神経細胞の反応は弱く、強いコントラストになると反応が強くなります。しかし、コントラストの増加に応じて神経細胞の応答が増加するコントラストの範囲(変化域)はかなり限られていて、この変化域の外ではコントラストが変化しても応答の大きさはほとんど変化しません。
しかしながら、私たちは広い範囲のコントラストの違いを検出することができます。強いコントラストの刺激を見ていれば強いコントラストの弁別ができるようになり、弱いコントラストの刺激を見ていれば弱いコントラストの弁別ができるようになります。このように広い範囲のコントラストの違いを区別するためには、神経細胞の変化域が周囲の平均コントラストに応じて移動する(順応)と好都合です。
研究手法と成果
研究チームでは、このようなコントラスト順応がヒトの脳の中のどこで起きているかを検討するため、機能的磁気共鳴画像法(fMRI)を用いてヒト被験者での実験を行いました。実験には、チェッカーボードパターンの白黒を15回反転させる視覚刺激を用いました(図B)。例えば、13%のコントラストの順応刺激を60秒間提示し続けた後、コントラストを4倍に増加した刺激を3秒間提示すると視覚野からの信号は一時的に増加しました(図C)。コントラストを4分の1に減弱した刺激を提示すると、V1野からV3野までの領野では、信号は一時的に減少しました(図C)。順応刺激を提示する前の信号のレベルを基準にして、いろいろなコントラストのテスト刺激により引き起こされた反応のピーク値をプロットすると、図Dの緑線のようにシグモイド関数に似た形のコントラストー応答曲線が得られました。
さて、順応刺激のコントラストを変化させ、例えば25%にすると、視覚野からの信号は一時的には上昇しますが、50秒程度の時間を経てほぼ元と同じレベルまで戻ってきました。この順応状態でいろいろなコントラストのテスト刺激を与え応答のピーク値をプロットすると、コントラストー応答曲線は右方向にほぼ並行に移動し、赤線のカーブが得られました(図D)。順応刺激のコントラストを減少させ、例えば6.3%にした場合には、コントラストー応答曲線は左方向へほぼ並行に移動し、青線のカーブが得られました(図D)。これらの結果は、順応によって、順応刺激のコントラストを中心とするコントラストの範囲に応答の変化域が動的に移動する、即ち、初めに予想した順応現象がV1野、V2野、V3野で実際に起こっていたことを示しました。
さらに、この研究では思いがけない発見がありました。より高次の領野であるV4野(図A)の反応においては、テスト刺激のコントラスト変化が増加であっても減少であっても信号は増加しました(図E)。
研究の意義
これまでの機能的磁気共鳴画像法による研究では、脳機能の時間的に変化しない側面を中心に調べられてきました。今回の研究では、独自の事象関連測定法を開発することにより、コントラストへの順応という脳機能の動的側面を精密に研究し、コントラストへの順応はV1野ですでに起こっていることを明らかにしました。さらに、V4野の神経細胞は変化したコントラストの内容ではなく、変化が起こったこと自体を検出する反応をしていることが示唆されました。
