




Background
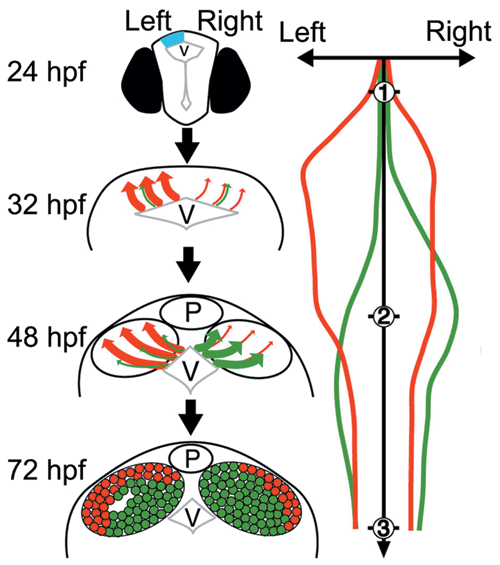
(Left column) Cartoons of frontal views of the dorsal diencephalon in the developing zebrafish brain. At 18-24 hours post-fertilization (hpf), Nodal and its downstream genes (light blue) are activated in the left dorsal diencephalon, and then neural precursors for the medial (green) and lateral subnuclei (red) are generated through stages 32 hpf and 48 hpf to form an asymmetric organization in the habenular subnuclei as observed at stage 72 hpf. (Right column) Schematic representation of developmental changes in the putative neurogenetic activities for the wild-type embryo. The green and red lines indicate the neurogenetic activities for the precursors of the medial and lateral subnuclei, respectively. The numbers on the vertical axis indicate developmental stage by the day post-fertilization. P, pineal organ; V, diencephalic ventricle * P: Pineal. V: Ventricle.
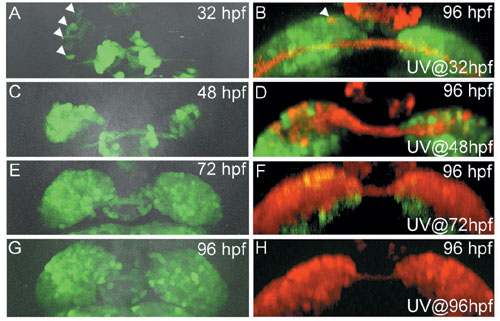
(A-H) Images of the transgenic fish showing the expression of HuC:Kaede during development (dorsal views; A, C, E, G) and the distribution at 96-hours post-fertilization (hpf) of cells which were photoconverted at specific stages (frontal views; B, D, F, H). Developmental stages and stages when we photoconverted Kaede are shown in the upper right and lower right corner of each panel, respectively. Arrowheads in (A) show asymmetric expression of non-photoconverted HuC:Kaede (green) in the left habenular anlage. Arrows in (B) and (D) show the photoconverted cells (red) in the lateral and medial subnuclei, respectively. More photoconverted cells were detected in the medial subnuclei by photoconversion at later stages (D, F, H). A bracket in (D) shows the habenular neuropils.
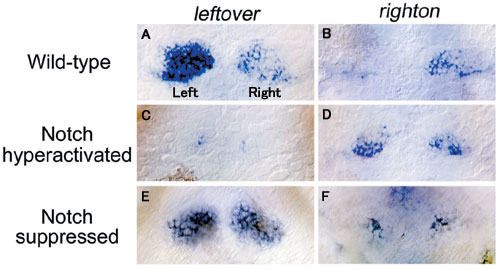
Dorsal views of wild-type (A, B) and genetically-manipulated embryos in which Notch signal is hyperactivated (C, D) or suppressed (E, F) showing leftover (A, C, E), righton (B, D, F) mRNA expression in the habenulae. leftover is expressed more in the left habenula (A) and righton is expressed more in the right habenula (B) in wild-type. Hyperactivation of Notch signal at 32 hpf lead to a decrease in leftover expression on both sides (C) with a large number of neurons expressing righton ectopically in the left habenula (D). Suppression of Notch signal leads to more leftover expression in the right habenula (E) and a corresponding decrease in righton expression (F).
Brain asymmetry is a conserved feature of the vertebrate nervous system and is thought to be an advantageous mechanism for efficiently processing information, since devoting one hemisphere to a particular function leaves the other hemisphere free to perform other functions. Recent studies have revealed asymmetry in the gene expression patterns and neural connections of the brain. However, the developmental mechanisms that contribute to the formation of these anatomical differences between both sides of the brain remains undetermined. Which developmental process, for example, is responsible for the establishment of left-right asymmetry? Our laboratory investigated this problem by analyzing development of the habenular nucleus and left-right asymmetry in the embryos of transgenic zebrafish.1)
Research Results
The habenula in zebrafish can be subdivided into medial and lateral subnuclei, each of which is composed of different types of neuron.2) We suspected that asymmetric development of the habenular subnuclei might occur because neurons of the medial and lateral subnuclei are born at different stages during development. This difference in the timing of neurogenesis may specify the types of neurons and the orientation of asymmetry. To address this hypothesis, we examined the birth dates of neurons in the medial and lateral subnuclei. We found that neural precursors for the lateral subnuclei were born at earlier stages than those for the medial subnuclei (Fig. 1). More neurons for the early-born lateral subnuclei were generated on the left side, while more neurons for the late-born medial subnuclei were generated on the right side (Fig. 1). Using Kaede, a protein that changes from a green to red fluorescence under UV irradiation, we traced the migration patterns of cells that were differentiated more on the left side and confirmed that early-born cells migrate to the lateral subnuclei (Fig. 2). Our results suggest that neuronal birth dates correlate with specificity and orientation of asymmetry in habenular subnuclei.
To determine how differential timing of neurogenesis specifies neural cell fate, we focused on Notch signaling that suppresses neurogenetic activity in the neural stem cells. Neurogenetic activity in embryos was genetically manipulated by hyperactivation or repression of Notch signaling during development, and the expression of different genes in the habenular subnuclei were analyzed (Fig. 3). When the hyperactivation of Notch signal prevented neurogenesis during early developmental stages, the lateral subnuclear precursors did not develop and the medial subnuclear precursor cells differentiated ectopically in the left habenulae (Fig 3-C, D). In the mutant which is defective in Notch signal, neurogenesis was promoted prematurely and the lateral subnuclear precursors were observed on both left and right sides (Fig. 3-E, F). These results suggest that timing of neurogenesis determines neuronal specificity for the medial and lateral habenular subnuclei, each of which shows left-right asymmetry.
Future Expectations
Left-right asymmetry in the brain may also determine some aspects of coordinated social behavior by regulating the direction (left-sided or right-sided) and the magnitude (how large the left-right difference is) of the asymmetry. In contrast to the digital-like selection mechanism between left and right sides, the developmental mechanism which we found seems to be an analog mechanism that controls the magnitude of the left-right differentiation. In the future, analyses using model animals such as the zebrafish should reveal the neural mechanism that controls adaptive behavior based on the left-right asymmetry of the brain.
