Introduction
Most of our movements depend on the skillful utilization of what our brains have learned and memorized over the course of our lives. The indispensable role played by the cerebellum was first identified more than ninety years ago by lesion effects on experimental animals and clinical studies on patients with cerebellar disorders. Around 1970, Drs. Marr, Ito (Special Advisor, RIKEN BSI) and Albus independently proposed that "optimal motion is learned within the cerebellar circuitry utilizing error signals that is produced during actual motion and mediated to the cerebellar cortex". Furthermore, Ito et al. (1982) discovered that the synapses on the cerebellar Purkinje cells have a plasticity that is called "long-term depression (LTD). For more than thirty years, this Marr-Ito-Albus hypothesis has been thoroughly examined and challenged. Representative controversial view against it is that the cerebellum contains the information required for motor learning but the learning itself occurs at its output destinations such as the cerebellar and vestibular nuclei (Miles and Lisberger, 1981). Our laboratory has investigated the implications of this issue over many years and now proposes that motor memory shifts transsynaptically.
Inter-Synaptic Shifting of Trace of Motor Learning
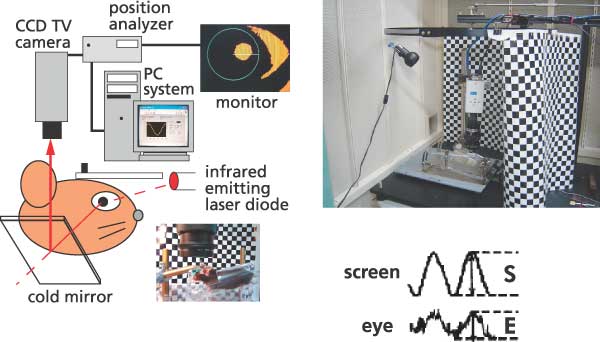
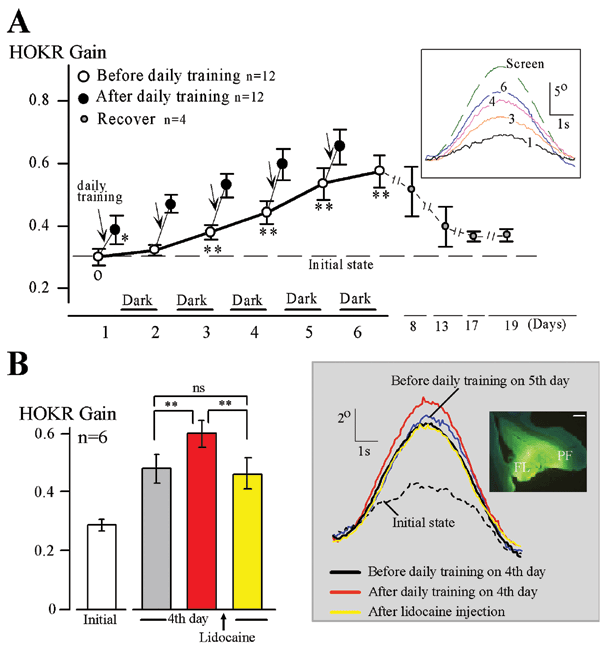
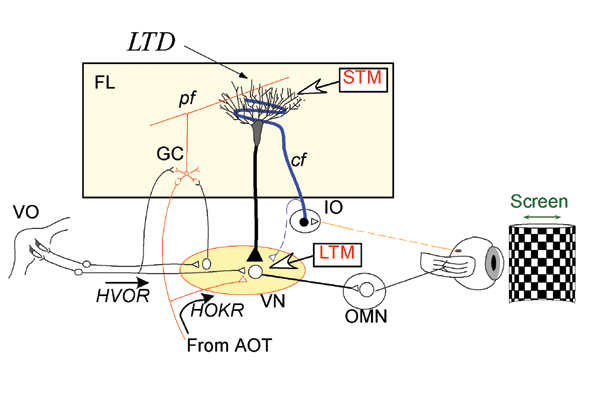
Our findings resulted from studies performed jointly with Dr. Shutoh (currently of the School of Comprehensive Human Sciences, Graduate Univ. of Tsukuba) in which we developed a learning paradigm of long-term adaptation of the horizontal optokinetic response (HOKR) of mice. When we watch the scenery from the window of a train, our eyeballs move with the moving landscape keeping the image sharp, not blurry. This action is called HOKR. We can simulate this behavior in the laboratory, by moving checkered-patterned screen in front of a mouse in a sine-wave direction. The HOKR effect shifts the eyes of the mouse in the same sine-wave form. If the screen is moved too rapidly, the eyes of the mouse cannot follow its movement because the efficiency of HOKR (the HOKR gain) is low at the beginning. However, the HOKR gain will improve with training and then the mouse's eyes will be able to follow the screen movement. When we returned the mouse to a dark cage after its initial training and tested the HOKR again on the next day, all gain increase disappeared. However, when we repeated the 1-hour training every day for a week, HOKR gain increase lasted for several weeks. This is a long-term learning effect. In addition, we also found that the learning effect obtained by the training of a day is extinguished immediately when the cerebellum is anesthetized, but long-term training is not affected at all. Hence, short-term motor memory after a day of training is retained in the cerebellum, but long-term memory is stored elsewhere. We also confirmed electrophysiologically that memory trace obtained from motor learning is actually present at the output destinations of the cerebellum after one week of learning. Based on our results, we concluded that the cerebellum initially obtains motor memory but this memory then shifts to the vestibular nuclei trans-synaptically as learning progresses and remains there. These findings extend the original Marr-Ito-Albus theory, and add the cerebellar cortical-nuclear circuitry as the storage mechanism of motor memory. The transfer of the memory trace probably was the source of controversy. In addition, in a joint study with Dr. Itohara, Head of the Laboratory for Behavioral Genetics, we found that LTD plays an important role in the transfer of memory to the vestibular nuclei.
Future Topics
With very simple behavioral experiments using mice, we were able to put an end to almost four decades of controversy about function of the cerebellum, and propose that memory acquired in the cerebellum shifts transsynaptically for long-term retention-thus challenging conventional wisdom in brain science. We have also confirmed that the transfer of the trace of motor learning occurs in the primates. At present, our laboratory is looking for the mechanism for the memory trace transfer in motor learning morphologically, genetically, and electrophysiologically. A transfer of the memory trace in the neural circuit has already been suggested for the declarative memory system in which the hippocampus plays the main role, but the essential processes are not yet known. The transfer of the memory trace seems important for memory retention, and finding the mechanism that enables that process will yield some important clues that will help solve the mystery of memory.