はじめに
私たちの動作の大部分は、日常生活の中で脳が学習し、記憶したものを巧みに利用することで行われています。運動に小脳が不可欠な働きをしていることは、小脳を損傷した動物の観察や、小脳障害の患者さんの臨床報告などから、20世紀の前半から知られていました。小脳の働きについて、Marr、伊藤(BSI特別顧問)とAlbusは、1970年前後にそれぞれ独立に、小脳の回路に可塑性があり、運動中に生じたエラーによって小脳皮質を通る信号が修正され、運動の学習が行われるという仮説を提案しました。さらに伊藤ら(1982)は、小脳皮質のプルキンエ細胞のシナプスに長期抑圧(LTD)という可塑性があることを発見しました。このMarr-Ito-Albus説は以後40年近く、世界中で検討されてきましたが、いくつかの反対仮説も提出されました。その代表は、小脳皮質は運動学習に必要な情報を伝えるが、学習が実際に起こるのは出力先の小脳核や前庭核であるという考え方(Miles & Lisberger, 1981)です。Marr-Ito-Albus説をめぐる論争は、Kandelらの有名な教科書にも詳しく紹介されています。私たちの研究チームはこの論争の原因を長年調べてきましたが、最近それに決着をつけるとともに、運動の記憶がシナプスを超えて移動するという新しい概念を確立することができました。
運動の記憶痕跡のシナプス間移動
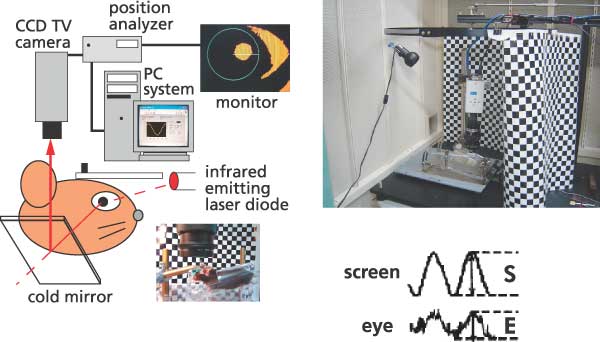
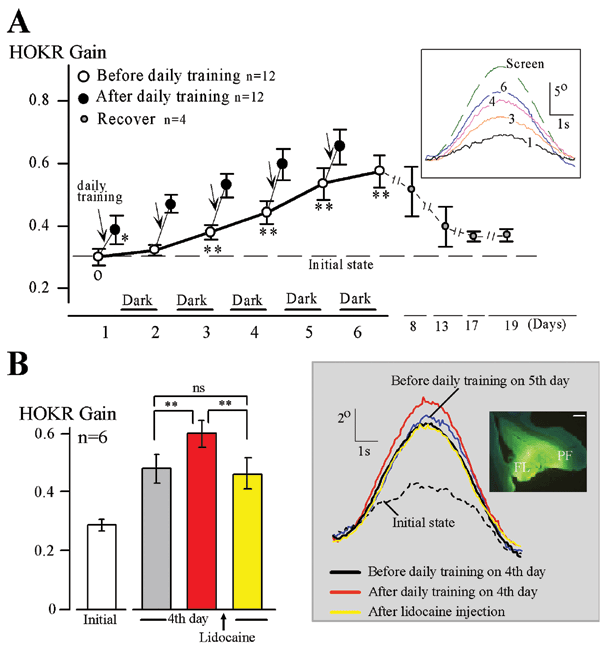
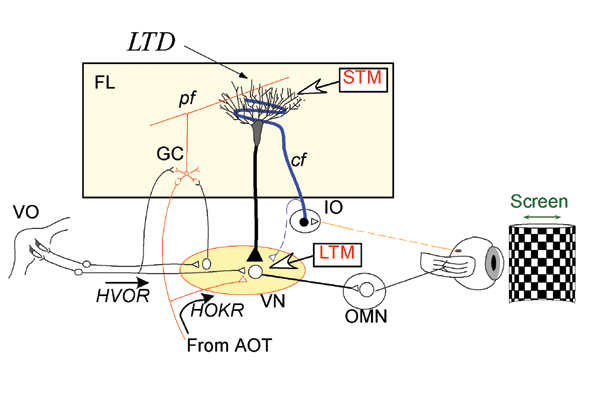
きっかけになったのは、首藤研究員(現筑波大学人間総合科学研究科)らとともに、マウスを使って水平性視機性眼球反応(HOKR)の長期適応の学習パラダイムをつくることに成功したことです。私たちが車窓から外の景色をぼんやりと眺めたときに、景色の動きに見合うだけ眼が動くことによって、景色がぶれずに見えるようにしているのが、HOKRという眼球運動です。実験室で、マウスの眼前にチェック模様のスクリーンを置き、それを正弦波状に動かすとHOKRによって眼も正弦波状に動きますが、スクリーンの動きを速めにしておきますと、最初はHOKRの効率が低いので、眼はスクリーンについてはいけません。しかし、マウスにスクリーンの動きを見る練習を1時間させると、学習が起こりHOKRの効率が向上し、スクリーンに眼が追従するようになります。練習終了後マウスをケージに戻して暗所飼育し翌日調べると、HOKRの効率は学習前に戻ってしまいますが、このような1時間の練習を1週間毎日繰り返すと、長期の学習が生じ、HOKRの効率が数週にわたって増加することを見つけました。さらに1日の練習で生じた学習の効果は、小脳皮質を局所麻酔剤で不活化すると直ちに消去されますが、1週間の練習で生じた学習の効果はまったく影響されないことに気づきました。つまり、1日の練習でできた運動学習の記憶は小脳皮質に維持されていますが、1週間の学習で長期化された運動学習の記憶は、もはや小脳皮質にはないということになります。さらに電気生理学的に、1週間の学習後には、小脳皮質の出力先の前庭核に運動学習の記憶痕跡が実際にあることを確かめました。このことから、運動学習の記憶は、Marr-Ito-Albus説のとおり、まず小脳皮質に獲得され、学習が進行するにつれて、シナプスを越えて前庭核に移動し、そこで固定化されることが分かりました。つまり、記憶の痕跡の移動が論争の原因の1つであったわけです。さらに、BSI行動遺伝学技術開発チームの糸原重美チームリーダーとの共同研究で、LTDが運動学習の記憶の獲得のみならず、前庭核への記憶の移動にも重要な役割を演じていることが分かりました。
これからの課題
マウスを用いた非常にシンプルな行動実験から、30年近く続いた小脳の役割をめぐる論争に決着をつけ、小脳皮質に獲得された記憶が、シナプスを越えて神経回路内を移動することにより長期間保持されるという、今までの脳科学の常識を覆すような考え方を提案することができました。現在、私の研究チームでは、運動の記憶痕跡の移動のメカニズムを、形態学、遺伝子発現、電気生理の手法で検討しています。また、運動の記憶痕跡の移動が、霊長類でも生じることを確かめました。記憶痕跡の神経回路内の移動は、海馬が主役を務める陳述記憶のシステムでも起こることが示唆されていますが、その本体はまったく分かっていません。記憶痕跡の移動は、記憶の保持にとって重要な意味を持つと思います。そのメカニズムを明らかにすることが、私たちの記憶の謎を解く重要な手がかりになることを期待しています。