




小脳皮質の神経回路は、主要な数種類の細胞がタイプごとにたくさん平行に並んだ単純な構造をしています。基礎回路の構成要素が少ないということは、そのひとつひとつの構成要素の数はとても多いということで、例えば小脳顆粒細胞は動物の脳の中で一番数の多い細胞となります。顆粒細胞の軸索は顆粒層からプルキンエ細胞とシナプスを形成していている分子層へ登ります。その軸索はT型に分岐し小脳表層を走行する平行線維となりプルキンエ細胞とen passantシナプスを形成します。齧歯動物の脳では、1つのプルキンエ細胞に対して~10,000,000,000もの顆粒細胞が~175,000のシナプス形成しており、各々のプルキンエ細胞はたいへんな数の顆粒細胞の影響を受けていることになります。この明らかにシンプルな構造は長年にわたり、小脳機能を解き明かしたいと願う研究者達を助長しました。おびただしい数の平行線維-プルキンエ細胞シナプスがメモリデバイスを構築していると考察され、そのアイデアはこれらシナプスの長期可塑性を示した実験により立証されました。そして平行線維の伝導速度が小さいために起こる顆粒細胞とプルキンエ細胞活動の間の時間差が、小脳機能を大きく左右するという説に至りました。後者の仮説は少なくとも概念上では、長期神経記憶とは区別されるであろう神経プロセシングです。運動失調と相関のある長期シナプス可塑性(長期抑制、LTD)で知られるミュータントマウスは数種ありますが、小脳において、シナプス長期可塑性の変化を伴わずに神経プロセシングの変化を示す実験データはとても少ないです。
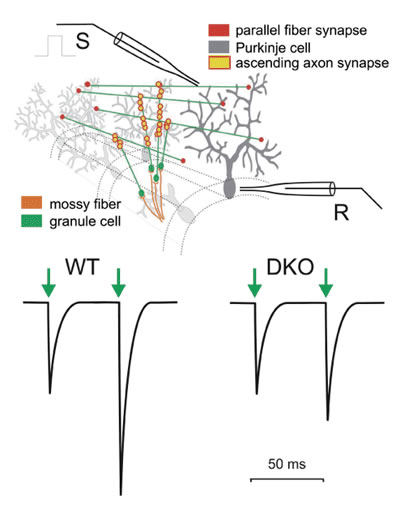
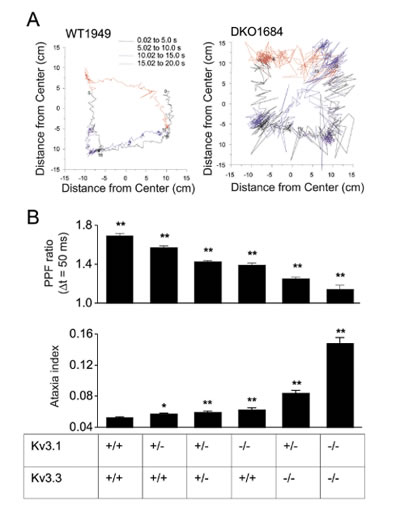
B) 機能的Kv3.1・Kv3.3の対立遺伝子を異なる数持つ欠損マウスより得られた短期可塑性(PPF ratio)と運動失調指標
そのため、私達はDr. Rolf. Joho(UT Southwestern, Dallas, USA)との共同研究で、小脳顆粒細胞に高発現してる2つのカリウムチャネル、Kv3.1とKv3.3の遺伝的欠損によって運動失調を示すマウスを同定したときは熱狂しました。私達はカリウムチャネル欠損マウスで見られる運動失調が、顆粒細胞-プルキンエ細胞シナプスにおける神経伝達を変化させることで生じているのではないかと仮説をたてました。これが事実ならば、この遺伝的欠損マウスの研究は小脳で生じる神経プロセシングの解明に役立つでしょう。私達はプルキンエ細胞のパッチクランプ記録を用いて、野生型マウスとカリウムチャネル欠損マウスの平行線維-プルキンエ細胞シナプスでの短期可塑性を調べました。野生型マウスでは平行線維の10~20活動電位毎にプルキンエ細胞で反応を生じますが、単一の活動電位ではプルキンエ細胞へのシナプスで伝達物質放出は生じにくくなります。しかし平行線維の束を刺激すると十分な数のシナプスが活性化されるので、プルキンエ細胞で大きく複合の興奮性後シナプス電流(EPSC)を記録することができます。次の活動電位がすぐに続くと神経伝達物質放出の確率が上がり、より大きいEPSCが観察されます。Paired- pulse-facilitationと呼ばれるこの現象は、最初のシナプス電流に対する次の電流の比(PPF ratio)として定量化されています。私達はKv3.1とKv3.3が欠損したダブルノックアウト(DKO)マウスでPPF ratioがたいへん小さいことを発見しました。(図1)第2のEPSCのサイズは前のEPSCに依存するので、PPFは短期可塑性のひとつの形です。
私達はそれから、非機能的Kv3.1・Kv3.3対立遺伝子の異なった数を持つマウスを調べました(図2-A)。運動能力はforce-plate actometerで定めた運動失調指標(Ataxia index)を使い定量化し(図2-B下)、対応するPPF ratio(図2-B上)と比較しました。.私達は運動失調指標とPPF ratioの両測定が遺伝子量に依存して変化することを発見しました。(図2)これらのデータは平行線維-プルキンエ細胞シナプスの短期可塑性と運動能力に関連があることを示します。しかし、重要な問題が2つ残っています。第一に、上述したデータは短期可塑性の変更と運動能力間の相関性のみであり因果関係を示すものではありません。この問題を解決するために現在Dr. Johoの研究室では、特定神経細胞種へのKv3チャネル機能の再導入されたマウスを作成しています。これらチャネルが顆粒細胞に再導入されることにより運動能力が改善されるならば、平行線維-プルキンエ細胞シナプスと運動能力の変化の間の因果関係が示されるでしょう。第2の問題は、平行線維全長にわたった顆粒細胞-プルキンエ細胞シナプス伝達の重要性についての古典的な討論であります。平行線維シナプスの大半がプルキンエ細胞を活性化させるには不十分であり、そしてプルキンエ細胞は主に顆粒細胞軸索の上行枝が形成するシナプスによって駆動されるという説や(図1)、分岐付近の平行線維のみが機能的なシナプスを形成するという説があります。それは、多くの平行線維シナプスが非機能的(silent synapses)であるとか、強力な抑制メカニズムが平行線維シナプスの効力を抑えていると討論されています。顆粒細胞が上行枝を介した局所的なプルキンエ細胞を活性化させるのか、それとも平行線維全長にそってより広く活性化させるのかを調べるために、私達は電位感受性色素と自家蛍光シグナルを使って実験を行ないました。苔状線維の白質刺激により平行線維の走行に沿ったビームのような蛍光強度変化が誘発されました。イメージングと細胞外記録を同時に行ない、ビームのような蛍光シグナルとプルキンエ細胞の活動(spiking)との関連を証明しました。これらのデータは、局所的に又は苔状線維-顆粒細胞経路に介して生じる平行線維の活性化が、平行線維の走行3mm以上にわたってシナプス後プルキンエ細胞を活性化させることができることを支持しています。
