





はじめに
脳機能ダイナミクス研究チーム(Laboratory for Human Brain Dynamics:LHBD)はヒトの脳機能を研究するため、非侵襲的手法を用いている。現在進行中のプロジェクトでは、単純な感覚刺激がどのように処理されるかを解明することに焦点を絞り、この情報を基に、感情および芸術鑑賞といった高次認知機能が、脳全体にわたってどのような神経活動で表現されているのかを探っている。
手法と背景
われわれ研究チームは、脳磁図(magnetoencephalography:MEG)を使用している。この手法は脳神経の電気的活動に伴って発生する微弱な磁場を検出する。脳からセンサーに至る短い距離を光速で移動するMEG信号を、頭部を覆う100箇所以上のセンサーで、ミリ秒単位、ときにはそれ以上の速さでも、計測することが可能である。同一刺激に対して示される非常に多様な反応を説明するため、多くの研究者は、ランダムな脳活動(random brain activity)という概念を受け入れ、「脳ノイズ」と名づけた。脳機能の「信号+ノイズ」モデルは、MEGから得られる加算平均データの平均値を正当化する論理的根拠となった。一方、われわれ研究チームは、大脳皮質、小脳、および脳幹での活動源をミリ秒単位で推定し、断層図に表示する技術、磁場断層撮影法(magnetic field tomography:MFT)を利用し、このような神経細胞活動の多様性の由来を観察してきた。MFTの結果により、背景となる脳活動と誘発反応との非線形結合を証明し、脳機能に関する「信号+ノイズ」という見解に対する最強の反証を示した。
(Moradi et al.、NeuroImage, 18:595-609, 2003)。
MEGでは信号源推定がどのくらいの精度で可能なのか?
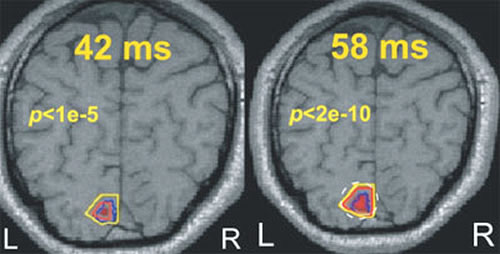
MEGにおける脳の信号源推定に関する問題は、コンピュータでシュミレーションしたMEGデータ、実データ、または多数の単純な感覚刺激を平均化した実データから、一つあるいは複数の点を信号源として、どの位良い精度で取り出されるかを検証することだと、通常言われている。しかし、1つまたは数個の信号発信源モデルでは生物学的実体は適切に反映されないので、大抵の場合このようなアプローチは無意味である。活動源推定の問題への取り組みを意味あるものとするため、われわれ研究チームは視覚系に関する知識と、認知機能表現研究チームの機能的MRI(functional MRI:fMRI)の専門知識を融合した。われわれのMFTの結果から得られる時間依存性統計パラメータマッピング(SPM)の画像とfMRIから得られる同様の画像が比較された。同一刺激と、同じ4人の被験者を使い、1次視覚野、VIの活動に驚くべき一致を明らかにすることができた。刺激開始後40ミリ秒近辺のpost-MFT SPMは、fMRI SPMの数ミリメーター以内であった(図1)。数ミリ秒以内でpost-MFT SPMは有線前野において広範な活動を示す。続いて、VIの活動の新しい波が、ちょうど50ミリ秒後に再び現われ、よく知られるピ-クに70ミリ秒で到達する。さらに研究を進め、小脳と脳幹における活動源推定を検証するため、ヒトの眼球運動過程に関する知識を利用することにより、正中線を越えて注視中枢を分離するのに十分な感度をMFTが有することを証明することができた。

初期感覚反応
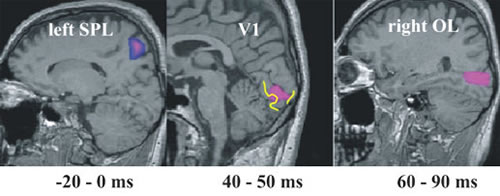
MEGデータの単一試行の正確な信号源推定は、外部刺激を処理における第1次視覚野の役割に関する新しい洞察が得られる。視覚システムにおいて、われわれは100ミリ秒以内の注意変調を検出した。刺激提示前に注意(attention)が必要となったとき、上頭頂小葉(SPL)の活動が亢進することが、 MFT分析により、確認された(図2)。刺激を除去したとき、fMRIの実験ではSPL活動が認められた。体性感覚システムにおいては、第1次体性感覚皮質, SIの時間依存性反応が確認された。最初の数百ミリ秒間に、SIでの活動は刺激特性によりもっぱら推進されるが、0.5秒後にはSIの活動は、異なった外部刺激が存在している間でさえも、被験者が刺激について考える内容を反映するようになる(図3)。聴覚系においては、左右の大脳半球における異なる聴覚感覚記憶痕跡に関連する単純な音刺激に対する反応において、明確な非対称性が確認された。このような側性効果は利き手および性別に依存する(Ioannides et al., NeuroImage, 19: 1061-1075, 2003)。
処理段階
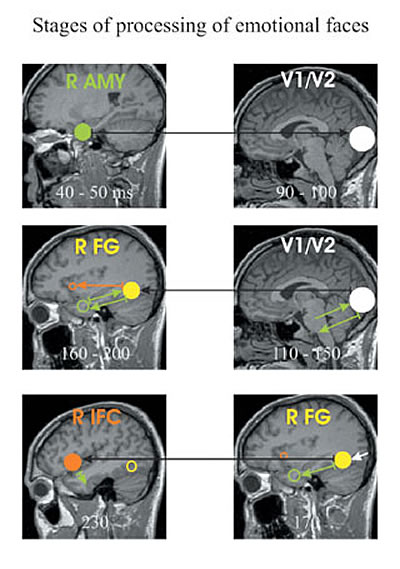
複雑な刺激に対する正常な脳の反応は、相互に影響しあう部位のネットーワークにおける処理段階(stages of processing)の観点から説明される。われわれは、感情を表現した表情の認識課題の処理段階について研究してきた。健常者では、微弱であるものの、ごく初期の扁桃体活動(50ミリ秒以内の)とVI(およそ80ミリ秒)間で相互作用は確立しているが、このような活動の連携は統合失調症の患者では認められない。より後期の活動を調べるときに、処理段階という概念は特に有用となる。健常者では、脳領野間の相互作用は、はっきりと処理段階としてグループ化され、順序だてて連続して行われる(図4)。統合失調症の患者では、同一課題は強く、限局された活動をもたらすが、領野間の連携が不充分なため孤立化し、完全な処理段階を形成していない。
夢と音楽
急速眼球運動(rapid eye movement:REM)すなわち「夢」睡眠時に記録されたMEGデータのMFTの結果、REMサッケード(衝動性眼球運動)の直前に現われる一連の活動を示した。眼窩前頭皮質に始まり、扁桃体、海馬傍回へと続き、眼球に対し運動開始の指令を出す脳幹に達するまで、連続して認められた。VIが活動を一瞬中断した後、サッケード開始後200ミリ秒近辺で再活動がはっきりと認められた。REM睡眠時には、VIの再活動に続いて左右扁桃体の再活動が出現する(Ioannides et al., Cerebral Cortex 14:56-72, 2004)。音楽実験に関しては、われわれは実際の音楽を使い、この目的のための特別な手法を開発することにより、楽譜の時間パターンと脳の活動部位を捕らえ、そして一致化を試みた(Popescu et al., NeuroImage 2004)。予想したとおり、結果は音楽が非常に広範囲の脳領域を活動させることを証明した。音楽刺激に対して初期段階の聴覚野は、他の聴覚刺激に対する場合と極めてよく似た反応を示す。数秒後には、さらに広範な活動が広い脳領野、特に小脳、運動野、および前頭葉に出現する。運動関連領野はリズムに反応するのに対し、前頭葉はより長時間スケールで活動する統合体として機能し、おそらく単純な音楽の断片を、より「意味のある」そして組織化された時間パターンに関連付けると思われる。web site(http://www.hbd.brain.riken.jp)にログインして、音楽に耳を傾け、音楽を鑑賞している時の脳活動のアニメーションをお楽しみ下さい。
共通のテーマは何か?
ただ1つのネットーワーク上の結び目をごく短時間眺めただけでは、MEGの信号およびリアルタイムの活動は、どの領野一つをとってみても、関連性に欠けるように見える。しかし、適切な方法で仔細に眺めると、統合された活動が、複雑な課題を極めて巧みに処理段階へと組織化する様子が見えてくる。そしてそこでは、与えられた課題に取り組むたびに、わずかに異なる領野が、わずかに異なる時間で補充されている。一部の病態においては、このような複雑な組織化が完全ではないと思われる。極端な反例をあげれば、われわれは夢を見たり、あるいは芸術を鑑賞するとき、これらの過程は一体となって絶妙なハーモニーを醸し出し、知覚、感情および意識といった統一された活動を生み出す。
脳機能ダイナミクス研究チームでは、共同研究の提案を募集しています。確かな行動結果が得られている実験設計が、プロジェクトとして理想的です。数少ない被験者を対象に、非常に時間がかかるデータ処理や分析手法を駆使して、脳の活動部位とタイミング、および活動部位の連携を、明らかにします。
